
Strouhaloniscellus biokovoensis, Bedek & Taiti, 2009
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2196.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:0308AF08-94B2-4C2F-AFAC-1058C640CB4A |
|
DOI |
https://doi.org/10.5281/zenodo.5333383 |
|
persistent identifier |
https://treatment.plazi.org/id/97263F35-3A3B-4B1E-B63D-F5D1DE82B312 |
|
taxon LSID |
lsid:zoobank.org:act:97263F35-3A3B-4B1E-B63D-F5D1DE82B312 |
|
treatment provided by |
Felipe |
|
scientific name |
Strouhaloniscellus biokovoensis |
| status |
sp. nov. |
Strouhaloniscellus biokovoensis View in CoL n. sp.
Material examined. Holotype. ♂ Croatia, Dalmatia , Mt Biokovo , Jama iznad Saranača, ca. 8 km ENE of Podgora, 4314'55.99''N, 1710'46.4''E ( WGS84 ), 760 m, 19.VIII.2007, leg. M. Lukić, CNHM IT1619 View Materials .
Paratypes. 4 ♀ ♀, same data as holotype, CNHM IT1055 View Materials ; 1 ♀, same data as holotype, MZUF 9298 View Materials ; 1 ♂, 1 ♀, same locality, 25.III.2003, leg. R . Ozimec , MZUF 9299 View Materials ; 1 ♀, same locality, 25. V .2004, leg. R. Ozimec , CNHM IT269 View Materials ; 1 ♀, same locality, 2.IV.2006, leg. J. Bedek, CNHM IT491 View Materials ; 3 ♀ ♀, same locality, 22. VI .2007, leg. M. Lukić, CNHM; 1 ♂, same locality, 19.VIII.2007, leg. J. Bedek, CNHM IT1054 View Materials .
Diagnosis. A species characterised by the small size (2.8 mm long), dorsum with a honeycombed basal structure, pereion with an oblique ridge on the epimera and 2+2 large tubercles on pereionites 1–6 and 1+1 on pereionite 7, male pereiopod 7 with a stout apically rounded seta on carpus, and male pleopod 1 exopod with a quadrangular distal part bent outwards.
Description. Maximum size: ♂, 2.8 x 0.8 mm; ♀, 2.8 x 1.2 mm.
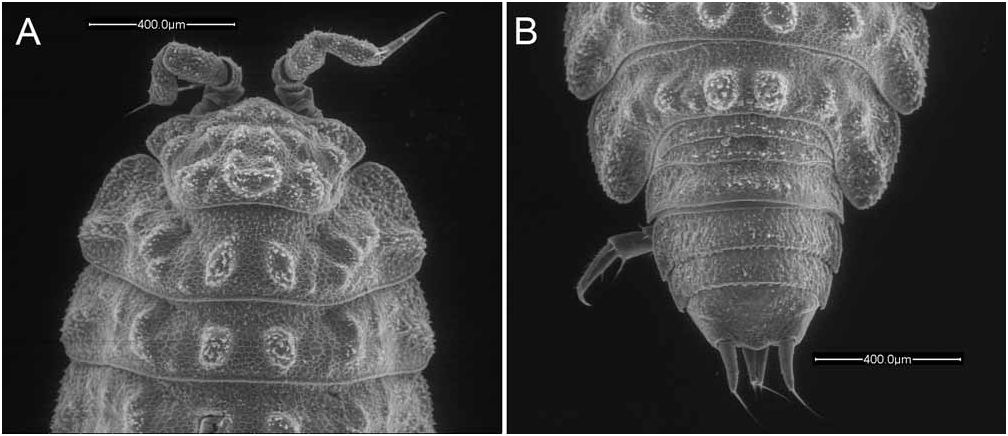
Body not strongly convex ( Figs 1A,B View FIGURE 1 , 2A View FIGURE 2 ). Dorsum with a honeycombed basal structure ( Fig. 1A,B View FIGURE 1 ) and some fan-like scale-setae ( Fig. 2B View FIGURE 2 ). Dorsal ornamentation present only on cephalon and pereion ( Figs 1A,B View FIGURE 1 , 2A View FIGURE 2 ): cephalon with three rows of tubercles on vertex, six tubercles in frontal row, six in middle row, and three in posterior row, middle one of which most prominent and horseshoe-shaped; pereionites 1–6 with an oblique ridge on epimera, two large tubercles and two to three additional very small tubercles per side; pereionite 7 with an oblique ridge on the epimeron, one large tubercle and four additional small tubercles per side. Cephalon ( Fig. 1A View FIGURE 1 ) with an obtuse median lobe and trapezoidal lateral lobes protruding at sides.
Pereionite 1 ( Figs 1A View FIGURE 1 , 2A View FIGURE 2 ) with rounded fronto-lateral corners and slightly sinuous posterior margin; pereionites 2–4 ( Fig. 2A View FIGURE 2 ) with almost straight posterior margin; pereionites 5–7 ( Fig. 2A View FIGURE 2 ) with epimera pointing backwards. Pleon ( Figs 1B View FIGURE 1 , 2C View FIGURE 2 ) much narrower than pereion; pleonites 3–5 with small epimera and short posterior points. Telson ( Figs 1B View FIGURE 1 , 2C View FIGURE 2 ) with distal part trapezoidal with slightly concave sides and broadly rounded apex.
Antennula ( Fig. 2D View FIGURE 2 ) of three articles; third article with a short spine and six to eight long aesthetascs at apex.
Antenna ( Fig. 2E View FIGURE 2 ) short and stout; fifth article of peduncle slightly longer than flagellum with distal seta about 2/3 as long as article; flagellum of four articles with a long aesthetasc on third article.
Mandibles ( Fig. 2F, G View FIGURE 2 ) with two penicils on left and one on right; molar process without penicils.
Maxillule ( Fig. 2H View FIGURE 2 ) with inner branch bearing three penicils at apex, inner one much larger than other two; outer branch with 10 teeth and two long and thin setose stems.
Maxilla ( Fig. 2I View FIGURE 2 ) apically setose with outer lobe much smaller and shorter than inner one; inner lobe with about 12 long stout setae.
Maxilliped ( Fig. 2J View FIGURE 2 ) endite narrow with a triangular penicil at apex; palp distally with four rounded lobes with long setae, outer margin with two groups of two-three setae, and basal article with two compound setae; basis with a rounded outer lobe protruding posteriorly and covered with long setae on the margin.
Uropod ( Fig. 2C View FIGURE 2 ) with exopod slightly shorter than endopod and more distally inserted; exopod with five long pointed setae and endopod with a single long seta at apex.
Male. Pereiopods 1 ( Fig. 3A View FIGURE 3 ) to 6 without distinct sexual modifications. Pereiopod 7 ( Fig. 3B View FIGURE 3 ) carpus sternal margin bearing a strong stout seta with a rounded recurved distal part and a thin seta at apex. Genital papilla ( Fig. 3C View FIGURE 3 ) fusiform with a segmented setose apical part. Pleopod 1 ( Fig. 3C View FIGURE 3 ) exopod about twice as long as wide, with a quadrangular distal part bent outwards and bearing a rounded lobe distinctly protruding medially; endopod of two articles subequal in length, second article narrow with rounded apex and a row of setae in the basal part. Pleopod 2 ( Fig. 3D View FIGURE 3 ) exopod trapezoidal with a large seta on outer margin and many short setae on inner margin; endopod biarticulated, with proximal article very short and distal article ending in a long thin point slightly bifid at apex.
Etymology. The name of the new species refers to Mt Biokovo where many caves are located, including the vertical one where these specimens were collected.
Remarks. Jama iznad Saranača is a small pit cave, 20 m deep. There are two different floors in the pit, the first one at a depth of 8 m in partial darkness and covered with decomposing organic material, the second, at the bottom of the pit, in total darkness with much less organic material. Both floors were equally investigated, but the specimens were collected under stones only on the second floor. Pieces of wet wood placed on the second floor did not attract any specimens. On the second floor the air temperature varied from 10.5 to 13.3°C, and relative humidity from 80 to 100%.
The genus Strouhaloniscellus was erected by Tabacaru (1993:67) to accommodate the species Haplophthalmus anophthalmus , described by Strouhal (1939) from one specimen collected in a small cave near Opservatorij, Mt Bjelašnica, Sarajevo, Bosnia and Herzegovina. According to Tabacaru the genus is characterised by the male pleopod 1 with the distal part of the exopod narrowed, bent outwards and bearing a lobe near the inner margin, and the male pleopod 2 with the distal part of the endopod thin and waved. The specimens from Mt Biokovo that we have examined show the same general shape of male pleopod 1 exopod, while the male pleopod 2 endopod is straight in its terminal part. This last character is most probably a specific rather than a generic character.
Strouhaloniscellus biokovoensis n. sp. differs from S. anophthalmus , as well as in the above mentioned shape of the male pleopod 2 endopod, in the following characters: smaller size (2.8 vs. 4.8 mm long), dorsal ornamentation on cephalon and pereion more developed with more numerous tubercles; the cephalic lateral lobes are more protruding laterally; the distal sternal margin of the male pereiopod 7 carpus has a strong seta with rounded apex bearing a thin seta, instead of a digitiform lobe; the male pleopod 1 exopod has a wider distal part and a larger rounded lobe which is protruding from outer margin, and the endopod has a thicker distal part with rounded instead of pointed apex. Moreover in S. biokovoensis the dorsum shows a honeycombed basal structure which, according to Strouhal (1939), is absent in S. anophthalmus .
The diagnosis of the genus Strouhaloniscellus proposed by Tabacaru (1993) is here emended according to the characters of the new species. The male pereiopod 7 carpus with a strong stout seta on distal sternal margin and the male pleopod 1 exopod with distal part narrower than basal, bent outwards and bearing a rounded lobe are the apomorphic traits that distinguish the genus within the subfamily Haplophthalminae . The small epimera of pleonites 3-5 may also be an apomorphic trait as a secondary reduction due to the small size. In other four species, Cyphoniscellus herzegowinensis ( Verhoeff, 1900) , Vardaroniscus tetraceratus Buturovič, 1955 , Bulgaroniscus gueorguievi Vandel, 1965 , and Vandeloniscellus bulgaricus ( Vandel, 1967) , there is a reduction of the epimera of the third pleonite. This character is considered to be a primitive condition by Vandel (1967) while according to Tabacaru (1993) this is a secondary reduction which occurred in parallel in the different species. In the other Haplophthalminae the epimera of pleonites 3–5 are well developed and there is no interruption between pereion and pleon. According to Dalens (1977) this character of fundamental importance in the subfamily precedes the formation of dorsal ornamentation. It is very difficult to distinguish the phylogenetic affinities within the subfamily Haplophthalminae which includes about 30 genera ( Tabacaru 1993, Andreev 2004, Schmalfuss et al. 2004, Karaman & Horvatović 2008), since most of the genera are defined according to the dorsal ornamentation (number and disposition of tergal ribs and tubercles), a character which can be heavily affected by homoplasy.
Most probably Strouhaloniscellus has a wider distribution in the Balkans since a third species, not yet described, is certainly present in Montenegro (I. Karaman pers. com.).


| CNHM |
Cincinnati Museum of Natural History |
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.