
Solanum adamantium Gouvea
|
publication ID |
https://doi.org/ 10.1600/036364419x15710776741521 |
|
DOI |
https://doi.org/10.5281/zenodo.6342841 |
|
persistent identifier |
https://treatment.plazi.org/id/CC16A766-2E2D-FFB4-7248-F8DC7961FAD0 |
|
treatment provided by |
Valdenar |
|
scientific name |
Solanum adamantium Gouvea |
| status |
|
Solanum adamantium Gouvea View in CoL ˆ, sp. nov. T
YPE: BRAZIL. Minas Gerais: Mun . Gr~ ao Mogol , próximo à Praia do Vau, trecho final da trilha que leva do fim da rua Hilário Marinho à Praia do Vau, crescendo em solo arenoso à margem da trilha, 16°35, 13̎S, 42°53, 32̎W, 667 m, Jan 2018 (fl, fr), Y.F Gouvˆea & I.F. Gouvea ˆ254 (holotype: BHCB! [ BHCB189841 ]; isotypes: RB, HUEFS).
This new species differs from S. thomasiifolium in its branched inflorescences (vs. unbranched inflorescences in S. thomasiifolium ), in its adaxial leaf surface indumentum composed exclusively of stellate-glandular trichomes (vs. adaxial leaf surface indumentum composed of both simple- and stellate-glandular trichomes in S. thomasiifolium ) and in its corolla completely lacking interpetalar tissue (vs. corollas with
conspicuous interpetalar tissue in S. thomasiifolium ); also differs from S. buddleiifolium in its branched inflorescences (vs. unbranched inflorescences in S. buddleiifolium ), in its erect fruiting pedicels that hold the fruits upward (vs. pendent fruits in S. buddleiifolium ).
Shrubs to slender treelets up to 4 m, erect, unbranched to few-branched when young, older plants moderately- to much-
branched. Young stems terete, pale green to greenish pale yellow, densely tomentose with hyaline to ochraceous stellateglandular trichomes, the epidermis not to barely visible, the trichomes sessile to short-stalked with multiseriate stalks up to 0.65 mm long, 2–4 cells wide, the rays 7–15, porrect to multiradiate, usually eglandular and 1-celled, 2–3-celled when glandular, sometimes with an early deciduous capitate glandular distal cell, the midpoint 3–4-celled, equal or shorter than the rays, the distal cell always glandular, usually persistent; stems moderately to densely armed with flattened prickles up to 1 cm long and to 3.3 mm wide at base, straight, golden to ferruginous; bark of older stems dark grey. Sympodial units plurifoliate, the leaves not geminate. Leaves simple, lobed, 5–19.5 cm long, 2–13 cm wide, the blade lanceolate, chartaceous, slightly discolorous, green to light green adaxially, lighter green abaxially; adaxial surface densely tomentose with hyaline to ochraceous stellate-glandular trichomes, always visible, the trichomes sessile to short-stalked with multiseriate stalks up to 0.5 mm long, 3–8 cells wide, the rays 5–20, antrorse to multiangulate, the eglandular rays usually longer and 1-celled, the glandular rays usually shorter and 2–3-celled, these often deciduous, the midpoints 3-celled, slightly shorter to slightly longer than the rays; the abaxial surface densely tomentose (a little denser than the adaxial) with the epidermis usually barely visible, the trichomes like those of the adaxial surface; sparsely armed along the midrib and the primary veins of both surfaces with straight, laterally compressed prickles reaching up to 6.2 mm long and to 0.9 mm wide at the base; primary veins 6–8 pairs; base slightly cordate to rounded or obtuse, strongly asymmetric to symmetric, not decurrent onto the petiole; margins irregularly lobed to nearly serrate, usually with secondary lobes, the lobes up to 2 cm long and to 3.5 cm wide at base, acute or less often rounded at the apex; apex acute, rarely narrowly rounded; petiole 0.7–5(–7.4) cm, densely tomentose with trichomes like those of the stem, sparsely to moderately armed. Inflorescence a branched cyme, erect, furcate, less often trifurcate (unbranched inflorescence may be occasionally produced among the branched ones on the same shoot), terminal, becoming internodal (lateral) by the subsequent sympodial unit’ s development, the axis densely tomentose with trichomes like those of the stem, unarmed to densely armed, the prickles straight; the peduncles (0.7) 1–3.7 cm long; the rachis (1.2) 1.7–9.4 cm long; with 8–60 flowers (4–30 per rachis), up to 6 flowers open at the same time (3 per rachis); pedicel insertion points 0–1.8 cm apart, becoming more closely spaced towards the inflorescence’ s apex; pedicels 4–8.4 mm long in open flowers, straight, articulated at base, unarmed or sparsely armed with minute prickles, densely tomentose with trichomes like those of the stem, the epidermis usually purple. Flowers 5-merous, the plants andromonoecious, producing hermaphroditic flowers (longstyled) and functionally male short-styled flowers, which vary in proportion (number of long- vs short-styled flowers) between inflorescences, with the proximal flowers usually longstyled and the distal ones short-styled. Calyx obconic, green to purple, unarmed, densely tomentose with the epidermis usually visible, the trichomes like those of the stems; the calyx tube 0.6–1.9 mm long, 1.9–2.5 mm in diameter at the widest point, the lobes 0.8–2 mm long, 0.7–1.5 mm wide at the base, triangular, the apices acuminate and usually slightly attenuate. Corolla (0.8) 1.5–2.2 cm in diameter, lilac to whitish lilac with the midvein purple, deeply stellate, slightly to markedly zygomorphic, with the three upper lobes closer to each other
than to the two lower ones, lobed 2/3 to 3/4 of its length, the sinuses deeper between the 3 upper and 2 lower lobes, interpetalar tissue absent, the tube 2–4.7 mm long, the lobes (3.1) 6–8 mm long, (1.8) 2.5–4 mm wide, narrowly triangular to nearly oblong or lanceolate, the apex acuminate to acute, densely to moderately tomentose abaxially with trichomes like those of the stems, glabrescent adaxially with some scattered trichomes near the apex and/or on the midvein. Stamens equal or slightly unequal, with 1–3 stamens slightly longer than the others; free portion of the filaments 1–2 mm long, glabrous; anthers (4.1)7.5– 6.5 mm long, 1–1.5 mm wide, 0.8–1.1 mm thick at the widest point, lanceolate, narrowed towards the apex, sagittate at base, connivent, poricidal at the tips, the pores about the same diameter as the anther apices, not lengthening with age, the epidermis smooth. Ovary cupuliform, 2-locular, densely pubescent with simple glandular trichomes mixed with some poorly developed, usually 1–3-rayed, stellate trichomes, these glandular or not; style 6.5–10 mm long in long-styled flowers, 1.2–2.3 mm long in short-styled flowers, cylindrical, sparsely to moderately puberulent with the same trichomes as those of the ovary; stigma clavate, up to 0.4 mm long in long-styled flowers, papillose, green when fresh. Fruit a globose berry, 7.2–9 mm long and 7.6–9.8 mm wide when mature, the pericarp smooth, marbled dark green and green when immature, reddish orange at maturity, moderately and evenly puberulent with short simple glandular trichomes all over the mature fruit surface; fruiting pedicels 5–7.3 mm long, unarmed or sparsely and minutely armed; fruiting calyx not accrescent, the tube 0.9–1.2 mm long and 3.9–5 mm in diameter at the widest point, the lobes 1.8–4 mm long, 1.2–2.1 mm wide at base. Seeds ca. 4–12 per berry, ca. 2.7–3.2 mm long, 3.4–4.3 mm wide, 0.9–1.5 mm thick, reniform, beige. Chromosome number: Not known. Figure 1 View FIG .
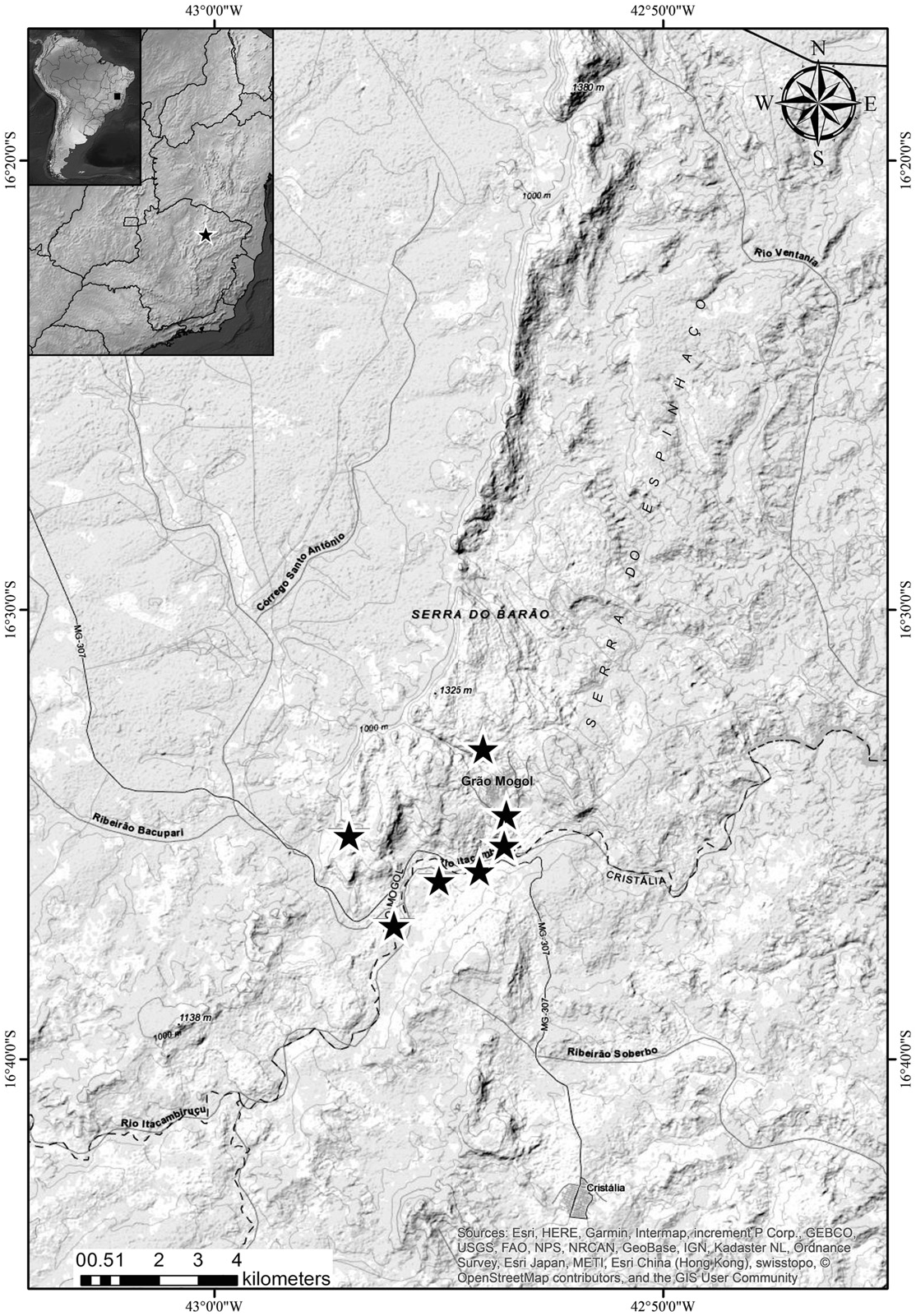
Distribution —This new species is endemic to the Gr~ ao Mogol range, part of the northern portion of the Minas Gerais State’ s Espinhaço range ( Fig. 2 View FIG ). Solanum adamantium occurs at elevations ranging from 650 to ca. 1000 m.
Ecology and Habitat — Solanum adamantium grows on and among rocks in high elevation quartzite/sandstone rocky fields (i.e. campo rupestre vegetation), and in nutrient-poor sandy soils associated with them, which also extend towards and along the lower lands of the Itacambiruçu river valley (See Pirani et al. 2003 for more details on the geography and the floristic composition of Gr~ ao Mogol’ s region). The campos rupestres are characterized, besides the strictly edaphic component, by extreme environmental conditions (i.e. hydric restriction, high solar radiation, wide daily temperature range, high elevation, and continuous exposure to winds; Giulietti et al. 1997; Rapini et al. 2008) to which this kind of vegetation is subjected. These conditions seem to have led to morphological convergences among the unrelated species inhabiting it (e.g. reduced and/or imbricate leaves usually restricted to the branch tips; see Giulietti et al. 1987), which are also observed in S. adamantium , S. buddleiifolium , and S. thomasiifolium . Like many ‘spiny’ Solanum species, however, S. adamantium can grow in, and colonize, somewhat disturbed and unstable sites, which is where several specimens were observed by the first author during fieldwork. Its flowers have morphologically adapted anthers for buzz pollination (i.e. anthers dehiscing by apical pores; Fig. 1E View FIG ) by vibrating bees, as in most Solanum species. The marked zygomorphy displayed by the corollas of older individuals of S. adamantium ( Fig. 1E View FIG ) is relatively rare in Solanum ( Knapp 2002; Bohs et al. 2007), especially among
Brazilian species. This unusual corolla shape (i.e. bilateral symmetry) may reflect a more specific plant-pollinator interaction (see Fenster et al. 2004), but it requires further study. The reddish orange colour of the S. adamantium fruits ( Fig. 1F View FIG ), together with the absence of scent and the permanent attachment suggest that its primary dispersal agents are birds (ornithochory; Van der Pijl 1972), which requires further investigation to be confirmed.
Phenology —Flowering specimens were found from December to October, indicating that S. adamantium may bloom and fruit year-round.
Etymology —The epithet adamantium is derived from the Latin “adamas,” meaning diamond; a reference to the history of the Minas Gerais State and the region of Gr~ ao Mogol (where the world’ s first extraction of diamonds from rocks was recorded; Karfunkel et al. 1994; Chaves et al. 2009), which are marked by the presence of this mineral.
Preliminary Conservation Status —We propose that if a formal conservation assessment were prepared and submitted to the IUCN, that this species would be classified as Endangered (EN) given its Extent of occurrence (EOO) of 24.5 km 2, and area of occupancy (AOO) of 24 km 2 ( IUCN 2017). The extremely narrow EOO of S. adamantium suggests that it might merit the status of CR. However, this is tempered by the incipient nature of our knowledge about this species (especially regarding the extent of its range), its ability to grow in somewhat disturbed sites, as well as the existence of the Parque Estadual de Gr~ ao Mogol.
Notes — Solanum adamantium shares ecological and morphological features with S. buddleiifolium and S. thomasiifolium , which in Stern et al. (2011) were resolved as sisters in one of the two lineages composing the poorly known Thomasiifolium clade. Both S. adamantium and S. buddleiifolium have narrower distributions, growing exclusively on rocks and in sandy soils of the elevated quartzite/sandstone rocky fields lying along the Espinhaço range. Solanum thomasiifolium , on the other hand, is more widely distributed, and also occurs (in a disjunct manner in relation to its Espinhaço range’ s populations) in the open vegetation of the sandy coastal lowlands (i.e. restinga; Araújo 1992). Despite sharing similar habitat preferences, these species were not found in sympatry thus far.
The morphological characters common to these three species seem to be related to the colonization of such extreme habitats (see Ecology and Habitat): small leaves restricted to the branch apices, short internodes, older stems with a nonphotosynthetic lignified bark up to the most proximal leaf, and densely indumented stems and leaves (see Figs. 1 View FIG , 3 View FIG ). These characteristics become more conspicuous with the plant’ s age, making species identification easier ( Figs. 1B View FIG , 3B View FIG ). Despite their overall similarity in the earlier stages of development, when in the reproductive phase, S. adamantium can be readily distinguished from S. buddleiifolium and S. thomasiifolium by having distinctive branched inflorescences rather than unbranched inflorescences (compare Figs. 1C, F View FIG , 3E, F View FIG ).
Solanum adamantium mostly resembles S. thomasiifolium , with which it shares glandular indumentum ( Figs. 1G View FIG , 3H View FIG ), somewhat cordate leaf bases, an erect inflorescence axis (even when fruiting), and erect fruiting pedicels holding the fruits upwards ( Figs. 1F View FIG , 3F View FIG ). However, in S. adamantium the leaves are not imbricate ( Fig. 1B View FIG ); the leaf blades are lanceolate, and covered exclusively by stellate-glandular trichomes ( Figs. 1B, G View FIG ); the corollas completely lack interpetalar tissue and are markedly zygomorphic ( Fig. 1E View FIG ); and the fruiting calyces
are inconspicuously accrescent. In S. thomasiifolium the leaves are imbricate with relatively short petioles, which give it a sessile appearance (mainly in older plants; Fig. 3F View FIG ); the leaf blades are cordate and covered in simple- and stellateglandular trichomes and simple-eglandular ones (the latter less conspicuous; Fig. 3H View FIG ); the corollas possess developed interpetalar tissue and are actinomorphic or slightly zygomorphic ( Fig. 3D View FIG ); and the fruiting calyces are conspicuously accrescent (up to ¾ of the fruit length).
Throughout the development of S. adamantium , the produced leaves gradually develop to a smaller size, making older specimens more similar to S. buddleiifolium in overall appearance ( Fig. 1B View FIG ). But S. adamantium can be recognized, in addition to the characters mentioned in the diagnosis, by its strongly glandular indumentum ( Fig. 1G View FIG ), while in S. buddleiifolium the indumentum is eglandular ( Fig. 3G View FIG ), or rarely with some glandular-stellate trichomes mixed with the eglandular-stellate trichomes, which are the vast majority.
Additional Specimens Examined— Brazil. — MINAS GERAIS: Mun. Gr~ ao Mogol, Córrego da Escurinha, campo arenoso, 16°35’S, 42°57’W, 750 m elev., 29 May 1988 (fl), A. Bidá et al. proj. C. F. C. R. 12102 ( RB!, SPF!); campo rupestre com abundancia ˆde arenito próximo de Grao~ Mogol, 12 April 1981 (fl), A. Furlan et al. proj. C. F. C. R. 750 ( RB!, SPF!); ca. 7 km à direçao~ sul da cidade, na estrada para Francisco Sá, campo rupestre/cerrado, afloramento de arenito e areia branca, 16°37’S, 42°56’W, 700–850 m elev., 15 October 1988 (fr), R. M. Harley et al. 25034 ( BM!); rua Hilário Marinho, campo rupestre cercado (loteamento) próximo ao fim da rua, crescendo em substrato acumulado sobre/entre rochas em campo rupestre onde houve supress~ ao da vegetaç~ ao, 16°34, 32̎ S, 42°53, 29̎ W, 785 m elev., January 2018 (fl, fr), Y. F. Gouvˆea & I. F. Gouvea ˆ255 ( BHCB!); interspread with wet rocky campo, ca. 8 km West of Gr~ ao Mogol, 950 m elev., 16 February 1969 (fl, fr), H. S. Irwin et al. 23368 ( HUEFS!, MO); Serra do Jambreiro, próximo ao Rio Itacambiruçu, 900 m elev., 10 May 1979 (fl, fr), H. C. de Lima et al. 995 ( BHCB!, JPB!, RB!); Trilha da Tropa, 16°33, 04̎ S, 42°54, 00̎ W, 990 m elev., 6 July 2007 (fl, fr), F. Marino et al. 288 ( BHCB!); escarpas à direita do Riacho Ribeir~ ao, 960 m elev., 21 May 1987 (fl), R. Mello-Silva & J. R. Pirani proj. C. F. C. R. 10772 ( BHCB!, HUEFS!); Vale do Rio Itacambiruçu, 4 September 1985 (fl, fr), R. Mello Silva et al. proj. C. F. C. R. 8342 ( BHCB!, SPF!); Jambreiro, margem direita da BR-251 (Gr~ ao Mogol – Montes Claros), área de extraç~ ao de quartzito, 687 m elev., 5 July 2011 (fl, fr), N. F. O Mota & C. N. Fraga 2275 ( BHCB!); Vale do rio Itacambiruçu, solo arenoso fino com rochas, ca. 16°36’S, 42°55’W, ca. 650 m elev., 12 December 1989 (fl, fr), J. R. Pirani et al. proj. C. F. C. R. 12563 ( BHCB!, SPF!); Estrada Grao~ Mogol – Cristália, ca. 6 km de Grao~ Mogol, campos rupestres com elementos de caatinga, 16°35, 47̎ S, 42°54, 05̎ W, ca. 650 m elev., 11 July 2001 (fl, fr), V. C. Souza et al.25675 ( ESA, RB!).


| BHCB |
Universidade Federal de Minas Gerais |
| RB |
Jardim Botânico do Rio de Janeiro |
| HUEFS |
Universidade Estadual de Feira de Santana |
| A |
Harvard University - Arnold Arboretum |
| C |
University of Copenhagen |
| F |
Field Museum of Natural History, Botany Department |
| R |
Departamento de Geologia, Universidad de Chile |
| SPF |
Universidade de São Paulo |
| M |
Botanische Staatssammlung München |
| BM |
Bristol Museum |
| S |
Department of Botany, Swedish Museum of Natural History |
| W |
Naturhistorisches Museum Wien |
| Y |
Yale University |
| I |
"Alexandru Ioan Cuza" University |
| H |
University of Helsinki |
| MO |
Missouri Botanical Garden |
| JPB |
Universidade Federal da Paraíba, Cidade Universitária |
| J |
University of the Witwatersrand |
| N |
Nanjing University |
| O |
Botanical Museum - University of Oslo |
| V |
Royal British Columbia Museum - Herbarium |
| ESA |
Universidade de São Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.