
Agonopterix ustjuzhanini, Buchner & Šumpich, 2020
|
publication ID |
https://doi.org/10.37520/aemnp.2020.013 |
|
publication LSID |
lsid:zoobank.org:pub:8B1A2630-FD4F-4592-9116-BAE629B235F4 |
|
persistent identifier |
https://treatment.plazi.org/id/CE1DC375-E32C-FFFC-EEC4-DED8FEF5BC9B |
|
treatment provided by |
Tatiana |
|
scientific name |
Agonopterix ustjuzhanini |
| status |
sp. nov. |
Agonopterix ustjuzhanini sp. nov.
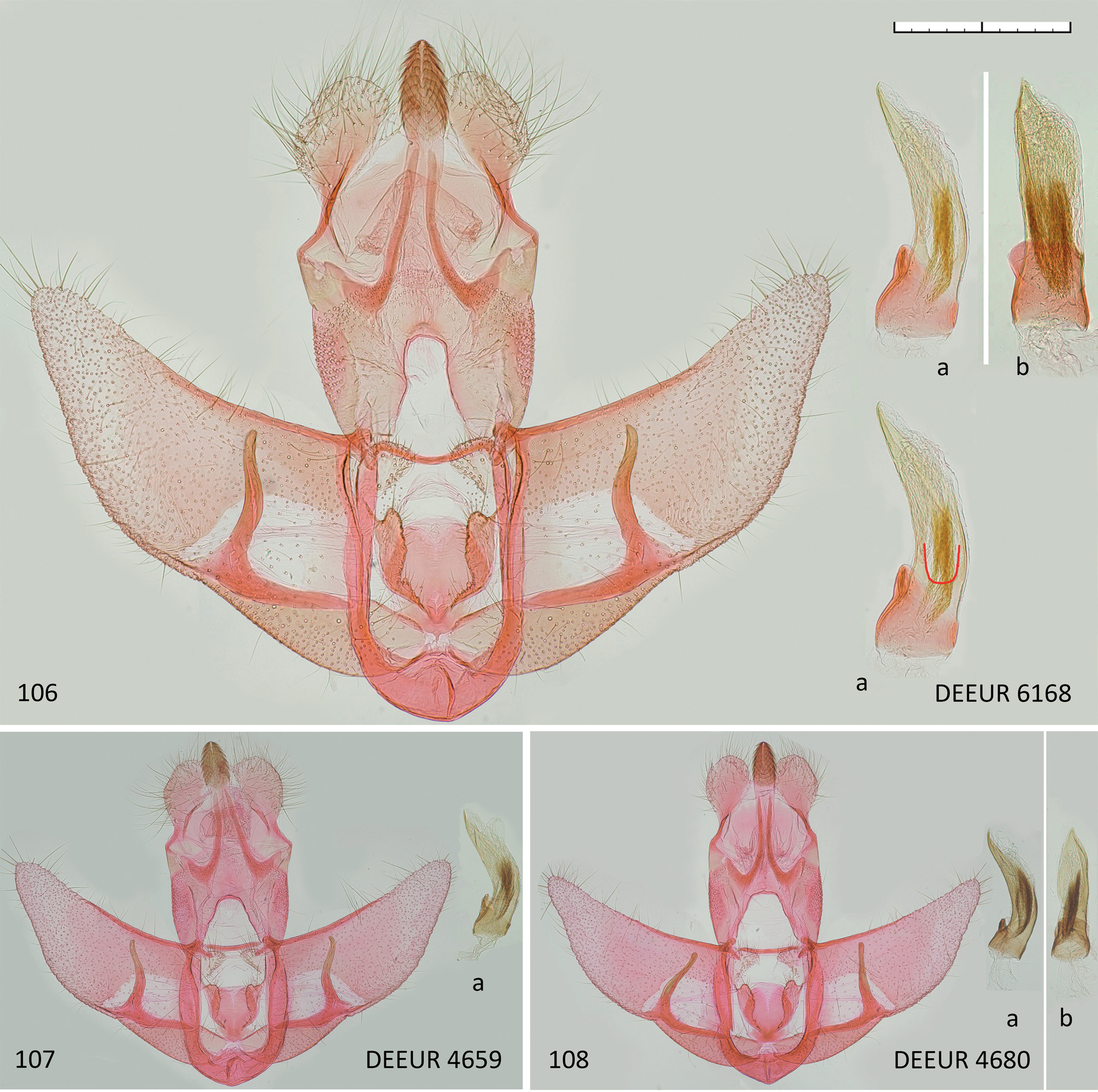
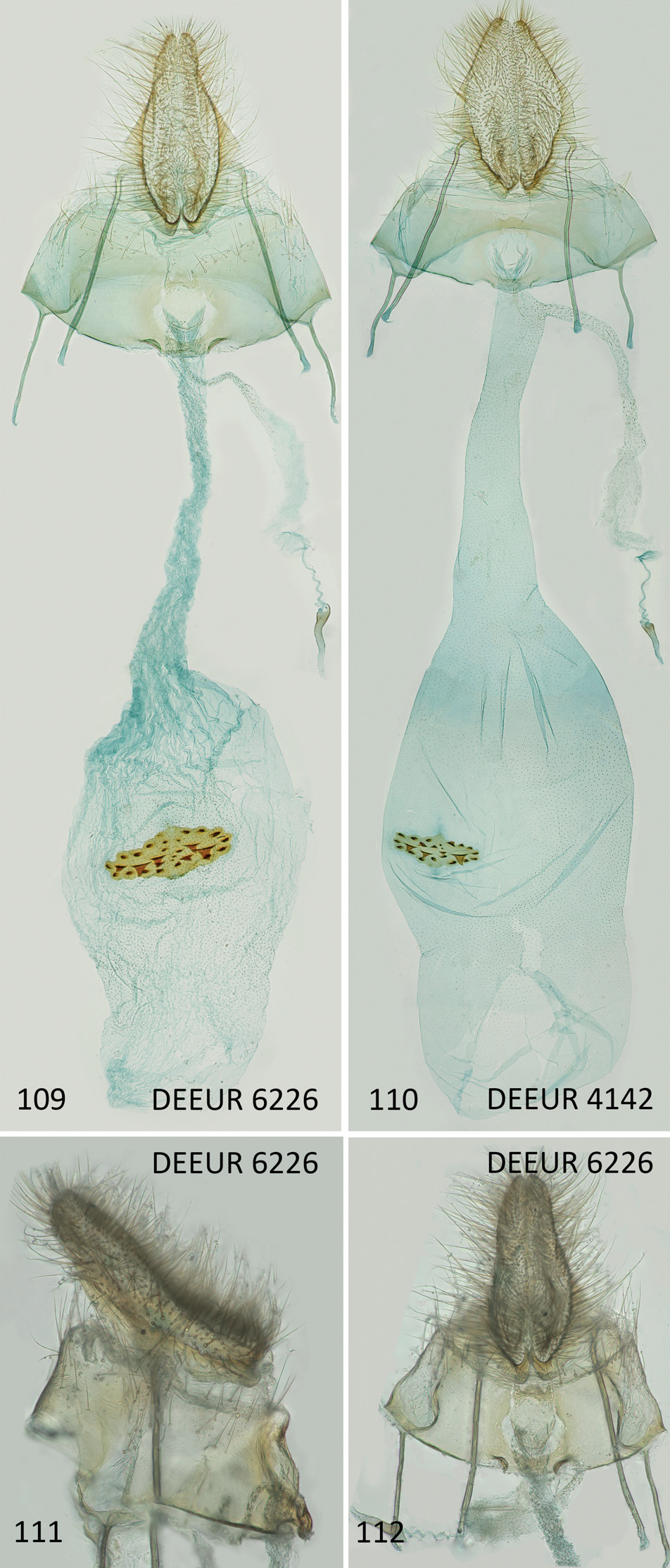
( Figs 99–112 View Figs 99–105 View Figs 106–108 View Figs 109–112 )
Type material. HOLOTYPE: Ƌ ( NMPC): RUSSIA: ALTAI REPUBLIC: Aktash vill., 50°19′12″N; 87°36′00″E, grassy steppe, rocks, 1400 m, 11.vii.2014, Barcode TLMF Lep 23279, gen. prep. DEEUR 6168, J. Šumpich leg. PARATYPES: 2 ƋƋ ( NMPC, PBSA):the same locality as holotype but M.Dvořák leg.; 1 Ƌ ( NMPC):the same locality as holotype but 29.vi.2014, M. Dvořák leg.; 1 ♀ ( NMPC): the same locality as holotype but 21.vi.2015, M. Dvořák leg. (gen. prep. DEEUR 6226 P. Buchner) (Barcode TLMF Lep 23318); Chulyshman valley, 45 km N of Ulagan vill., 51°01′03″N; 88°00′39″E, grassy steppe, rocks, 600 m, 27.–28. vi.2015, 1 Ƌ (Barcode NMPC-LEP-0159), J. Šumpich leg. ( NMPC); 1 Ƌ, the same locality but 4.–5.vii.2019, J. Šumpich leg. ( NMPC); Belyashi (Dzhazator) env. ( 56 km SE), rocky steppe and mountain meadows near Tara river, 2400 m, 49°39′45″N, 88°14′28″E, 25.–26.vii.2017, 4 ƋƋ (Barcode NMPC-LEP-0158), J. Šumpich leg. ( NMPC); Shebalino environs, 8 km W of Cherga village, 51.56° N, 85.48° E, 8.–9.vii.2019, 1 ♀, J. Šumpich leg. ( NMPC); Tabozhok valley, 10 km NE Kosh-Agach vill., Kurai Mts. Range, 2400 m, 50°05′N, 88°45′E, 2.viii.2016, 1 Ƌ ( TLMF Lep 21240), P. Huemer & B. Wiesmair leg. ( TLMF); Kosh-Agach distr., Ukok Plateau, northern part, 2400 m, 49°30′N, 88°06′E, 5.viii.2016, 1 Ƌ ( KLM Lep 06357), Ch. Wieser leg. ( LMK). BURYATIA: Hamar Daban Mts., Murtoy River near Gusinoe Ozero village, 51.2° N, 106.18° E, 19.vi.2002, 1 Ƌ ( TLMF Lep 19213), gen. prep. DEEUR 4659, K. Nupponen leg. ( RCKN). KAZAKHSTAN: Kokpek, 43.45° N, 78.66° E, 1.x 1994, 1 ♀ (Barcode NGS-29197-E01), gen. prep. DEEUR 4142, Gy. Fabian leg. ( RCGF); Katutau Mts., Konyrolen River, 44.21° N, 79.33° E, 2.x.2015, 1 Ƌ (Barcode TLMF Lep 19326), gen. prep. DEEUR 4680, K. Nupponen leg. ( RCKN).
Description. Adult ( Figs 99–105 View Figs 99–105 ). Wingspan 24.0– 27.5 mm. Head grey with brownish tinge, scales tipped whitish, face grey. Labial palp grey, segment 2 paler on inner side, with brownish tinge ventrally and speckled dark grey on outer side, segment 3 with partial dark grey ring at two-thirds and tip. Antenna fuscous. Thorax pale grey, becoming grey-brown in anterior third, tegulae pale grey. Forewing grey with brown tinge, with scattering of dark fuscous scales; basal field pale grey, edged by broad dark fuscous fascia in dorsal two-thirds, costa with blackish dot at base, costal spots blackish, extending along whole length of costa with tendency to be smaller and more clearly contrasting against ground colour in basal half; terminal spots blackish; oblique dots blackish, another at end of cell, consisting of white centre with blackish scales entirely surrounding it or these blackish scales weaker to absent on proximal side; diffuse spot on costal margin of cell dark grey, of variable size, sometimes indistinct and confluent with costal spots; plical dot indistinct; fringes pale greyish brown, with two fringe lines, distal one indistinct. Hindwing greyish brown, a little darker towards termen, with a few dark grey spots between veins, fringes similar to ground colour with two or three indistinct fringe lines. Abdomen grey.
Variation. We examined one specimen with grey or brownish grey colour entirely replaced by warm yellowish brown ( Fig. 101 View Figs 99–105 ).
Male genitalia ( Figs 106–108 View Figs 106–108 ). Socii medium-sized, broadly elliptic, 0.4–0.5 mm wide and 0.6–0.7 mm long, uncus triangular, indistinct, gnathos elliptic, medium-sized, 0.25–0.30 mm wide and 0.45–0.55 mm long, overtopping socii in standard preparation by about third to half of its length. Transtilla narrow, not widened medially, transtilla lobes semi-elliptic, 0.3 mm wide and long, gap between them 0.15–0.20 mm. Anellus medium-sized, round, length/ breadth 0.6/ 0.6 mm, gap to transtilla 0.3–0.4 mm, caudal margin with distinct V-shaped central incision filled with thin membrane, anellus lobes medium-sized, narrow elliptic, with distinct bulge towards transtilla which gives them semicordate appearance, length 0.5–0.6 mm, width 0.15–0.20 mm. Valva about 2.7–3.0 mm long, 1.2–1.4 mm broad at base, tapering to about 1.0 mm at origin of cuiller, then widening a little before evenly tapering to its rounded tip. Cuiller rather thin, 0.1 mm diameter in basal half, tapering to 0.05–0.07 mm in distal half, slightly S-shaped ( Fig. 107 View Figs 106–108 ) or sometimes only slightly bent outward ( Fig. 108 View Figs 106–108 ), ending shortly (about 0.1 mm) before costa, blunt.Aedeagus 1.3–1.4 mm long, bent (40–60°) in lateral view, diameter 0.3 mm, sclerotised basal parts with total length of about 0.4 mm, free section about 0.15 mm, in lateral view (basal process and concave edge must be on left side!) with distinct, U-shaped area paler than the rest, exceeding end of free section of basal sclerotisation ( Fig. 106 View Figs 106–108 , highlighted in the copy of aedeagus in lateral view, important feature in male genitalia to separate from similar species, details under diagnosis). In ventral view about as wide as in lateral view, but somewhat expanded at about 3/5 from base before tapering to sharp tip, free part of basal sclerotisation about as broad as aedeagus and with convex ending. Vesica (uneverted) with numerous tiny cornuti in 2 or 3 not clearly separable groups of about 1/2 of aedeagus length.
Female genitalia ( Figs 109–112 View Figs 109–112 ). Papilla analis about 1.5 mm long and 0.4 mm broad in lateral and 0.6 mm in ventral view (refers to unembedded genitalia), posterior apophysis 1.6 mm. Sternite VIII 0.8 mm long, maximum width 2.3 mm, anterior apophysis 0.6–0.7 mm.
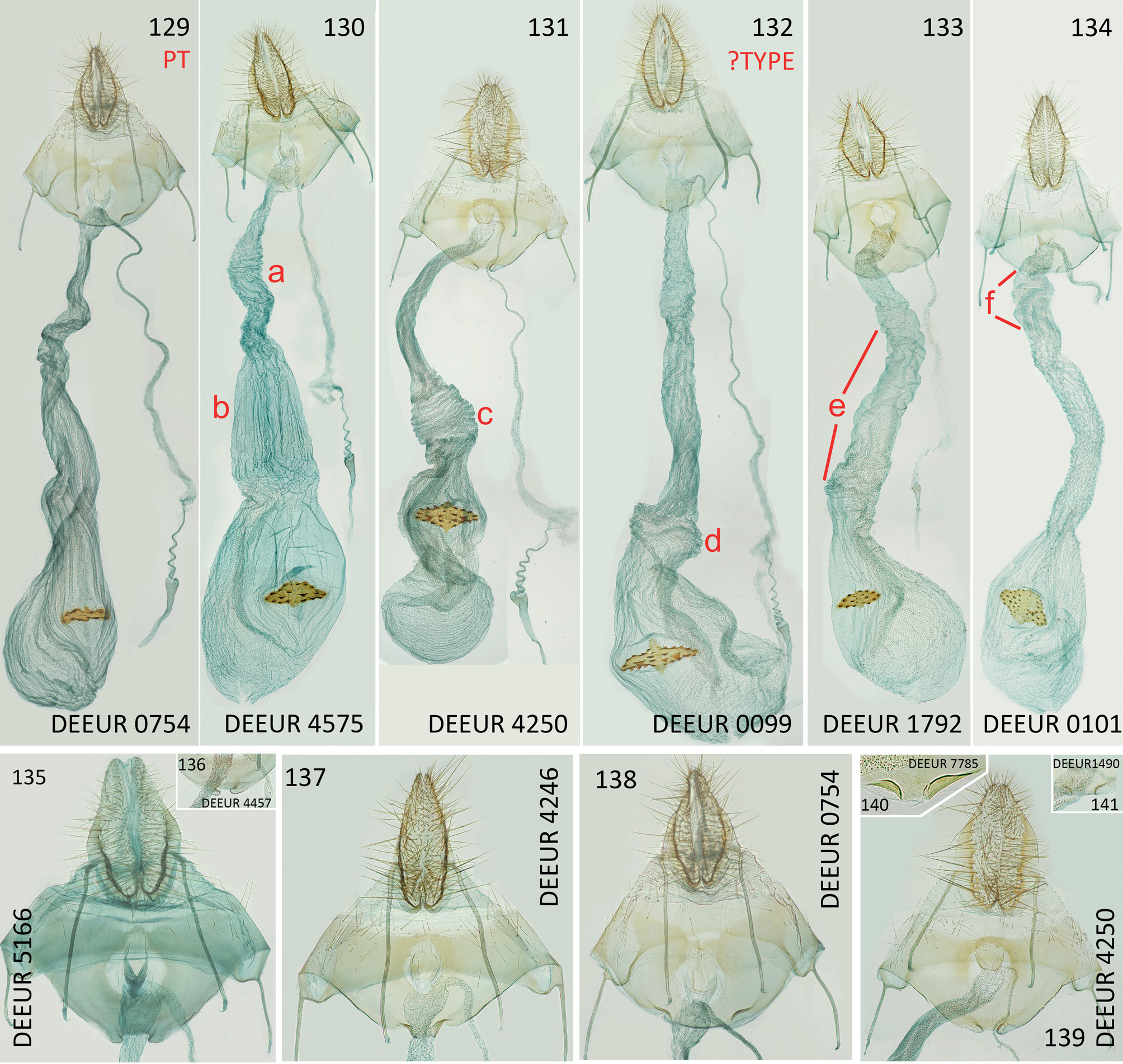
Distal edge of sternite VIII medially with U-shaped excavation of about two-thirds of ostium width, proximal edge of sternite VIII evenly concave, with 0.15–0.20 mm long fold on either side, distance between inner ends of folds 0.30–0.35 mm, ostium slightly proximal to middle of sternite VIII, round with diameter of about 0.3 mm, ductus bursae about 2.5–3.0 mm long, corpus bursae elliptic, 3.0–4.0 mm long and transverse diameter up to 2.0 mm, signum medium-sized with width/length ratio of about 0.7–1.0/ 0.3–0.4 mm, about 15–20 small teeth at its margin and about 4–6 larger, triangular teeth along transverse axis. Origin of ductus spermathecae 0.2 mm from ostium and ending with about 4 turns, ductus and corpus bursae covered with numerous tiny dots, apart from this without specifically distinct constant structures. Details of ductus bursae reflect life history, especially whether the specimen has mated or not, rather than specific features, which makes a detailed description useless (compare Figs 109–110 View Figs 109–112 ).
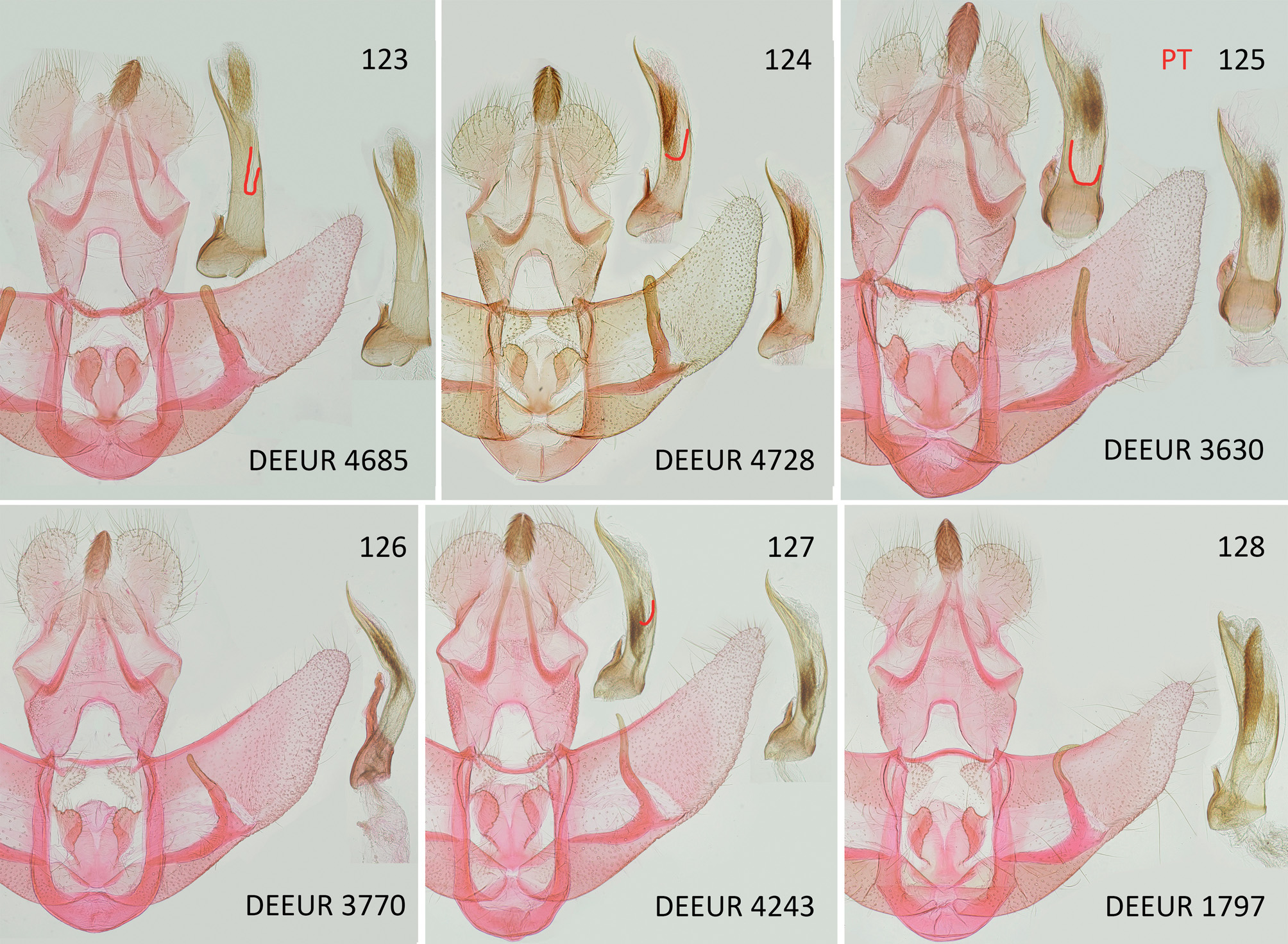
Differential diagnosis. Externally, the new species shares the basic wing patterns mainly with Agonopterix adspersella (Kollar, 1832) and A. thapsiella (Zeller, 1847) , and other closely related species, namely A. leucadensis (Rebel, 1932) , A. cervariella (Constant, 1884) , A. olusatri Corley & Buchner, 2019 , and A. cadurciella (Chrétien, 1914) . With wingspan up to 27.5 mm the average size exceeds all other species, but for determination this is of limited use because there is an overlap between 24 and 26 mm with other species; in addition, exceptionally small specimens can be found in every species. The new species is most similar to A. adspersella / A. thapsiella (these two species are treated as a species pair here, because without information about larval features and food plant it is not possible to determine every specimen of this pair to species level; it is not the intention of this paper to solve this problem, although some details are discussed further below). Reddish elements, especially in oblique dots and central dot with reddish brown instead of blackish scales, are often developed in A. adspersella / A. thapsiella but have not been found in A. ustjuzhanini . Besides this, many specimens of this very variable species pair are separable from A. ustjuzhanini only by dissection. In A. leucadensis , white scales in the central dot are usually completely absent, but they are rarely present as in A. ustjuzhanini , providing an overlap in external features in occasional specimens. However, with distribution of A. leucadensis restricted to Greece, as far as known, the risk of confusing these two species is low. In A. cervariella , the third segment of labial palp is pale flesh-red without black elements except for a few black scales at the very tip, and forewings show a flesh-red tinge, which helps to separate A. cervariella from all the other species mentioned here. In A. olusatri , the white scales in central dot are usually absent or only a few if present, and the pale colour of the basal field does not extend along costa, while in A. ustjuzhanini it tends to continue along the basal half of costa. Agonopterix cadurciella is comparatively easily separable externally by forewings with very pale ground colour, covered with diffuse, medium brown patches, best seen in Figs 113 and 115 View Figs 113–122 , but also in this species intraspecific variability makes it impossible to determine every specimen based only on external appearance. One example is a specimen ( Fig. 114 View Figs 113–122 ) with brown areas very extended and the interspersed black scales more numerous than usual. Another is a specimen ( Fig. 116 View Figs 113–122 ) where features usually distinct for A. cadurciella are nearly absent, and only the fact that it was reared from Seseli cf. tortuosum L. indicated that it could be A. cadurciella , which was confirmed by dissection.
In male genitalia, the new species differs from all the compared species in distinctly narrower socii and anellus lobes. Similar shape of cuiller is found in A. cadurciella only, but in this species it is much longer and clearly exceeds the costa of valva. The extent of pale area on the left side of aedeagus is a good feature to distinguish the new species from A. adspersella / thapsiella, where it is much smaller, and from A. olusatri , where it is clearly larger. Agonopterix leucadensis is best distinguished by the shape of cuiller which is unique in this group by being sharply hooked at its tip, and A. cervariella by the shape of aedeagus in lateral view, which is also unique in this group by being remarkably narrow with a sharp angle in its middle and the free section of basal sclerotisation reaching this bend.
Diagnosis based on female genitalia is shown in Table 2. Molecular data. BIN BOLD: ADC2512 (n = 10, 0 public, 6 from Altai). The average intraspecific divergence of the barcode region is 0.32% (maximum 0.80%)
Etymology. The species name is dedicated to Petr Ustjuzhanin ( Novosibirsk, Russia), an expert on Pterophoridae , who was helpful to the second author during his trips to Russia.
Biology. Food plant unknown so far, but with all species of the A. adspersella group feeding on Apiaceae , it is likely the same with A. ustjuzhanini .
Distribution. Russia ( Buryatia, the Altai Republic), Kazakhstan (Kokpek, Katutau Mts.).
Remarks. In the past, there was some confusion between related taxa of the A. adspersella group to which A. ustjuzhanini belongs. The “ Agonopterix adspersella group” can be understood in different ways: as a group of very closely related species comprising A. adspersella , A. thapsiella and A. chironiella (Constant, 1893) , but also as a broader group consisting of the species listed below. Specimens belonging to the first narrow group ( A. adspersella group sensu stricto) cannot always be determined to species level if no information about the preimaginal stages is available. Specimens of the broader group ( A. adspersella group sensu lato) may be very similar and undeterminable externally, but with genitalia dissection or barcoding, a safe determination is possible. Anyway, during identification all the species of the broader group should be kept in mind, and we include them for comparison.
A detailed treatment of species not found in the Altai Mts. in a paper dealing with the fauna of this region may be puzzling. But the description of A. ustjuzhanini brought to light so many doubtful and incorrect information about the A. adspersella group in actual literature that it was impossible to present a sufficient differential diagnosis without a closer look at this group.Also, we realised that features so far used for determination show a geographical variation and therefore it is essential to expand the look on the whole range of these species, which in this case inevitably leads far away from the Altai Mts. Finally, essential features for species delimination are found in preimaginal stages, especially the colour of thoracic legs of mature larvae ( Figs 142–145 View Figs 142–145 ). Information about corresponding larval and imaginal features (both external and in genitalia) is insufficient, which makes it difficult to impossible to discern adult features which represent only intraspecific variability from features which are of diagnostic value, thus leaving a considerable number of adults that lack information about larval features undeterminable at the moment. An additional reason to present larval features is motivation to fill this gap in the future.
Comments on the taxonomic position of problematic taxa from the Agonopterix adspersella group
Agonopterix adspersella ( Figs 117 View Figs 113–122 , 124 View Figs 123–128 , 131 View Figs 129–141 , 142 View Figs 142–145 ) was described in 1832 by Vincenz Kollar from “Mödling und Baden”, an area south of Vienna in Austria, characterised
by limestone hills and xerothermic climate. No information is given about food plant and appearance of the larva.
In NHMW there are several specimens under A. adspersella , matching the original description and collected in the type area. One of them may be the type specimen, although it is not quite clear. It has four labels, first is “Podevin”, a collector from 19th century whose collection includes many specimens from other lepidopterists, second is “coll. Zell.”, third is “ adspersella Koll. , Type ” handwritten by Rebel, fourth is “ adspersella Präparat 1” handwritten by Josef Klimesch. There is no further label data, neither date nor place. Klimesch reared specimens from larvae collected in 1935 in subalpine areas of “Grosser Pyhrgas”, Upper Austria, on Athamanta cretensis . To clarify whether these specimens belong to A. amanthicella (Heinemann, 1870) , reared from “ Bupleurum foliatum ” [sic!] and Meum athamanticum Jacq. from the Bavarian Alps, or to A. adspersella (Kollar, 1832) , he dissected the type specimen from NHMW. The investigation revealed complete agreement with the genitalia of A. adspersella , and the specimens reared from Athamanta cretensis and A. amanthicella were subsequently synonymised with A. adspersella ( KLIMESCH 1942). In the type area, larvae of A. adspersella are most often found on Bupleurum falcatum L.; further food plants there are Laserpitium siler L. and Seseli libanotis (L.) W.D.J.Koch. With the first pair of thoracic legs black, they match the description of larvae of A. feruliphila .
Agonopterix thapsiella (Zeller, 1847) ( Figs 123 View Figs 123–128 , 137 View Figs 129–141 , 144 View Figs 142–145 ) was described by Philipp Christoph Zeller from Sicily; larvae were collected from Thapsia garganica and a detailed description of the larva was given already by Zeller. The most important detail here is as follows: “Vorderbeine gelblich, braunfleckig” [= thoracic legs yellowish, with brown dots].
Agonopterix feruliphila (Millière, 1866) was described by Pierre Millière from Hyères ( France, Var); larvae were collected from Ferula nodiflora Pall. and a detailed description of the larva is given. The most important detail here is “Les pattes antérieures sont testacées avec le dernier article noir et luisant; les dix autres pattes sont unicolores et la couronne est jaunâtre” [= thoracic legs are yellowish brown with the last segment black and shiny; the other ten legs are unicolorous and the crown yellowish]. Agonopterix feruliphila was synonymised with A. thapsiella by REBEL (1901). Investigation of larval features ( RYMARCZYK et al. 2013) revealed that this conspecificity was incorrect. There are several differences in larvae reared from Thapsia garganica and those reared from Ferula nodiflora , most importantly in the colour of the first pair of thoracic legs, matching with the original descriptions of A. thapsiella and A. feruliphila . We fully agree with the conclusion that A. feruliphila is not a synonym of A. thapsiella . In the work of RYMARCZYK et al. (2013), genitalia of A. feruliphila and A. thapsiella are also compared, and features for determination of not-reared specimens are proposed. We agree that some specimens can be determined as one or the other species without a doubt; however, there remain specimens where even dissection does not bring a clear result.
There is a wide field of open questions for the future. The range of food plants is not yet known, and there is also the possibility that larval features might be modified by different chemistry of food plant. Without a safe tool to identify each specimen it is impossible to discern spe- cies-specific genitalia differences. Such an investigation must include the whole distribution range of A. adspersella / A. thapsiella , not only Europe, but also North Africa and the Asian part of its range. Until such investigations bring more clarity regarding this species pair, it is recommended to list doubtful specimens as A. adspersella / A. thapsiella . Agonopterix chironiella (Constant, 1893) ( Figs 119 View Figs 113–122 , 132 View Figs 129–141 , 145 View Figs 142–145 ) was described by Alexandre Constant from Alpes Maritimes ( France); larvae were collected from Opopanax chironium W. D. J. Koch and a description of the larva is given, but without detailed information on the colour of thoracic legs. Imagines are not directly compared with other species, only the remark “J`estime qu`elle trouvera sa place naturelle prés de Depressaria applana ” [= I think it will find its place near Depressaria applana – actually valid as A. heracliana (Linnaeus, 1758) ], with an additional remark that the new species is distinct enough to exclude the risk of confusion. Agonopterix thapsiella is not mentioned at all. For a long time this species was only known from France and Italy. In 2019, it was found in Greece for the first time: “ Peloponnes, Kalavryta, 800m, leg. larva 29.v.2019 from Opopanax chironium R. Seliger , e.p. 26.vi.2019 (RCRS, det. R. Seliger, conf. P. Buchner)”. Although this species is externally distinct and usually safely discernable from A. thapsiella , larval and genital features (and also barcode) show it is very close to A. thapsiella . It must be kept in mind that worn out specimens may be inseparable from A. adspersella / A. thapsiella .
In the past, the name A. chironiella was used for an undescribed species, now valid as A. olusatri , so any determination or record of A. chironiella from outside of its known range may belong to A. olusatri . However, the unexpected discovery of A. chironiella in Greece shows it is likely to be more widespread.
Agonopterix cadurciella (Chrétien, 1914) ( Figs 113-116 View Figs 113–122 , 127 View Figs 123–128 , 130, 135-136 View Figs 129–141 ) was described by Pierre Chrétien from Causse de Gramat ( France, Pyrenées). According to the description, larvae were collected from Hyoseris L. ( Asteraceae ) but it is likely that the food plant was misidentified: “Trois sujets obtenus en juillet de chenilles trouvées en juin, dans le Causse de Gramat, sur une Compositée défleurie, dont je n`ai pas gardé d`échantillon, mais qui m`a paru être un Hyoseris ” [= Three moths obtained in July from caterpillars found in June, in the Causse de Gramat, on a withered plant, of which I did not keep a sample, but which appeared to be a Hyoseris .]. In our opinion Hyoseris ( Asteraceae ) is surely not the food plant. Also HANNEMANN (1958) had doubts about this Asteraceae association, because all species from the A. adspersella group feed on Apiaceae . Although A. cadurciella is distinct and usually determinable externally at first glance (comparison in Figs 113–116 View Figs 113–122 ), its existence has been widely ignored. From all museums which were visited by the first author, beside MNHN, where the types are stored, only in TLMF a correctly identified specimen of A. cadurciella was found. In many other museums the specimens are either mixed with A. adspersella or with A. thapsiella or even with both (e.g. in NHMUK). So the misinterpretation of A. cadurciella as A. adspersella by RYMARCZYK et al. (2013) is understandable.
The specimens of A. adspersella , stored in NHMW and collected in its type area south of Vienna, clearly contradict the opinion of RYMARCZYK et al. (2013). There is an evidence for A. adspersella and A. cadurciella not being conspecific. In and around the type locality of A. adspersella there has never been found even a single specimen of A. cadurciella . The first author, who lives in this area and knows the microlepidoptera fauna very well, has never seen it there. Kollar described this species in 1832, so it is certainly possible that in nearly 200 years the fauna could have changed. Therefore, specimens of A. cadurciella were looked for in museum collections collected in Austria in the 19 th century. But although the majority of the large collections in Europe have been checked, no specimen of this species, collected in or near the type locality of A. adspersella or indeed anywhere in Austria could be found. On the other hand, the species with larva depicted as Fig. 142 View Figs 142–145 , reared from Bupleurum falcatum and corresponding in the black first pair of thoracic legs with A. feruliphila , is common in this area. This makes it most likely that Kollar, when describing A. adspersella , had in front of him specimens conspecific with the species which was described in 1866 by Millière as A. feruliphila , but not with the species which was described in 1914 by Chrétien as A. cadurciella .
In conclusion: A. feruliphila again becomes a synonym, but actually of A. adspersella , and not of A. thapsiella as was mentioned so far. This does not affect the specific difference between A. adspersella (= A. feruliphila ) and A. thapsiella , published by RYMARCZYK et al (2013) on the basis of differences in larval features.
Checklist of taxa from the Agonopterix adspersella
group
Agonopterix adspersella (Kollar, 1832) ( Haemylis) ; TL: Austria (near Vienna)
= amanthicella Heinemann, 1870 ( Depressaria ); TL: Europe (south) = athamanticella Steudel & Hofman, 1882 (? Agonopterix ) (mis- -spelling) = sabulatella Turati, 1921 ( Depressaria ); TL: Italy (Rome: Monte Autore) = karmeliella Amsel, 1935 ( Depressaria ); TL: “ Palestine ” = rubripunctella Amsel, 1935 ( Depressaria ); TL: “ Palestine ” = feruliphila Millière, 1866 ( Depressaria ) syn. nov.; TL: France
Agonopterix cadurciella (Chrétien, 1914) ( Depressaria ); TL: France ( Midi-Pyrénées: Causse de Gramat)
Agonopterix cervariella (Constant, 1884) ( Depressaria ); TL: France (central)
Agonopterix chironiella (Constant, 1893) ( Depressaria ); TL: France (Alpes Maritimes)
Agonopterix leucadensis (Rebel, 1932) ( Depressaria ); TL: Greece
Agonopterix olusatri Corley & Buchner, 2019 ; TL: Portugal
= chironiella auct., nec. Constant ( Agonopterix )
Agonopterix thapsiella (Zeller, 1847) ( Depressaria ); TL: Sicily
= linolotella Chrétien, 1929 (? Agonopterix ); TL: France
In some internet sources Depressaria reichlini Heinemann, 1870 is erroneously listed as a synonym of A. adspersella , but it actually belongs to D. pimpinellae Zeller, 1839 .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.