
Cribrinopsis, Carlgren, 1921
|
publication ID |
https://doi.org/ 10.1080/00222930600703532 |
|
persistent identifier |
https://treatment.plazi.org/id/CF17AF78-FFBA-3876-FDC3-B651FB19CA05 |
|
treatment provided by |
Felipe |
|
scientific name |
Cribrinopsis |
| status |
|
Genus Cribrinopsis View in CoL
Diagnosis (modified from Carlgren 1949)
Actiniidae with well-developed pedal disc. Column with adhesive verrucae or without these. Pseudospherules absent or present. Fosse distinct. Sphincter strong, circumscribed. Tentacles short, thick, their longitudinal muscles principally mesogloeal. Radial muscles of oral disc meso-ectodermal to mesogloeal. Numerous perfect mesenteries usually decamerously arranged. Well-developed mesenterial muscles. Gonads on mesenteries of the first cycle and on the other stronger mesenteries. Same number of mesenteries proximally and distally, or mesenteries slightly more numerous proximally. Size ranges (lengths) of large basitrichs in actinopharynx and tentacles always overlap. Cnidom: spirocysts, basitrichs, microbasic p-mastigophores A, and microbasic pmastigophores B.
Cribrinopsis albopunctata sp. nov.
( Figures 1 View Figure 1 , 2 View Figure 2 , 12 View Figure 12 )
Tealia lofotensis: Hand 1955, p 80 View in CoL ; Sebens and Laakso 1977, p 162.
Not Madoniactis lofotensis Danielssen 1890, p 47 View in CoL .
Material examined
Holotype: East Kamchatka: Avacha Bay, Starichkov Island , 13 m, 5 June 2003 ( KBPIG 261 /3) . Paratypes: East Kamchatka: Avacha Bay, Starichkov Island , 13 m, 5 June 2003, one specimen ( KBPIG 262 /4) ; 11 m, 18 June 2003, four specimens ( KBPIG 263 /5) .
Other specimens. East Kamchatka: Avacha Bay, Starichkov Island , 20 m, 9 September 2003, two specimens ( KBPIG 267 /9); Bezymyannaya Bay , 10 m, 1 October 2004, four specimens ( KBPIG 264 /6); November 2004, one specimen ( KBPIG 265 /7); Tri Brata Islands , 21 September 2002, 12– 13 m, two specimens ( KBPIG 266 /8). Commander Islands : Medny Island , intertidal, 4 August 1995, one specimen ( KBPIG 11 /1) .
Description
External structure. Known living specimens are not large, the height of the column and the diameter of the base rarely exceeding 7 cm. The largest contracted formalin-preserved specimen is about 4.5 cm in diameter, and 3 cm in height. The pedal disk is wide and strongly adhesive. In most specimens the cylindrical column is dark red or vermilion, sometimes paler, rose coloured or white, becoming paler toward the base. White adhesive verrucae are arranged in longitudinal rows corresponding to the exocoels and endocoels from the margin to the limbus, they become smaller toward the base. The most distal verrucae are larger, sometimes partly fused together. Verrucae over the endocoels may be larger and form longer rows than those over the exocoels, and in smaller specimens exocoel verrucae may appear only near the margin ( Figure 12A View Figure 12 ). Verrucae are not strongly adhesive, but may attach particles of gravel and broken shells. Occasionally marginal verrucae have a shallow dark depression in the centre, giving an impression of a perforation, but true perforated pseudospherules are not present. There is a deep fosse and short capitulum.
The oral disk is flat and circular, commonly of the same ground colour as the column, or sometimes differently coloured, greenish or olive. Dark red radial bands running from the middle of the oral disk to the fosse, outline the tentacle bases and form a characteristic colour pattern ( Figure 12B, D View Figure 12 ). In the pale or white specimens the disk may be uniformly coloured.
The tentacles, arranged decamerously in four cycles on the outer half of the oral disk, are not numerous, the largest examined specimen had 72 tentacles and another smaller specimen had 76 tentacles. They are conical, pointed and slightly longer than the radius of the oral disk, and in preserved specimens longitudinally striated, thick, and short. In life the tentacles are uniformly coloured red, rose, or white along most of their length, usually with the white patch on the bases.
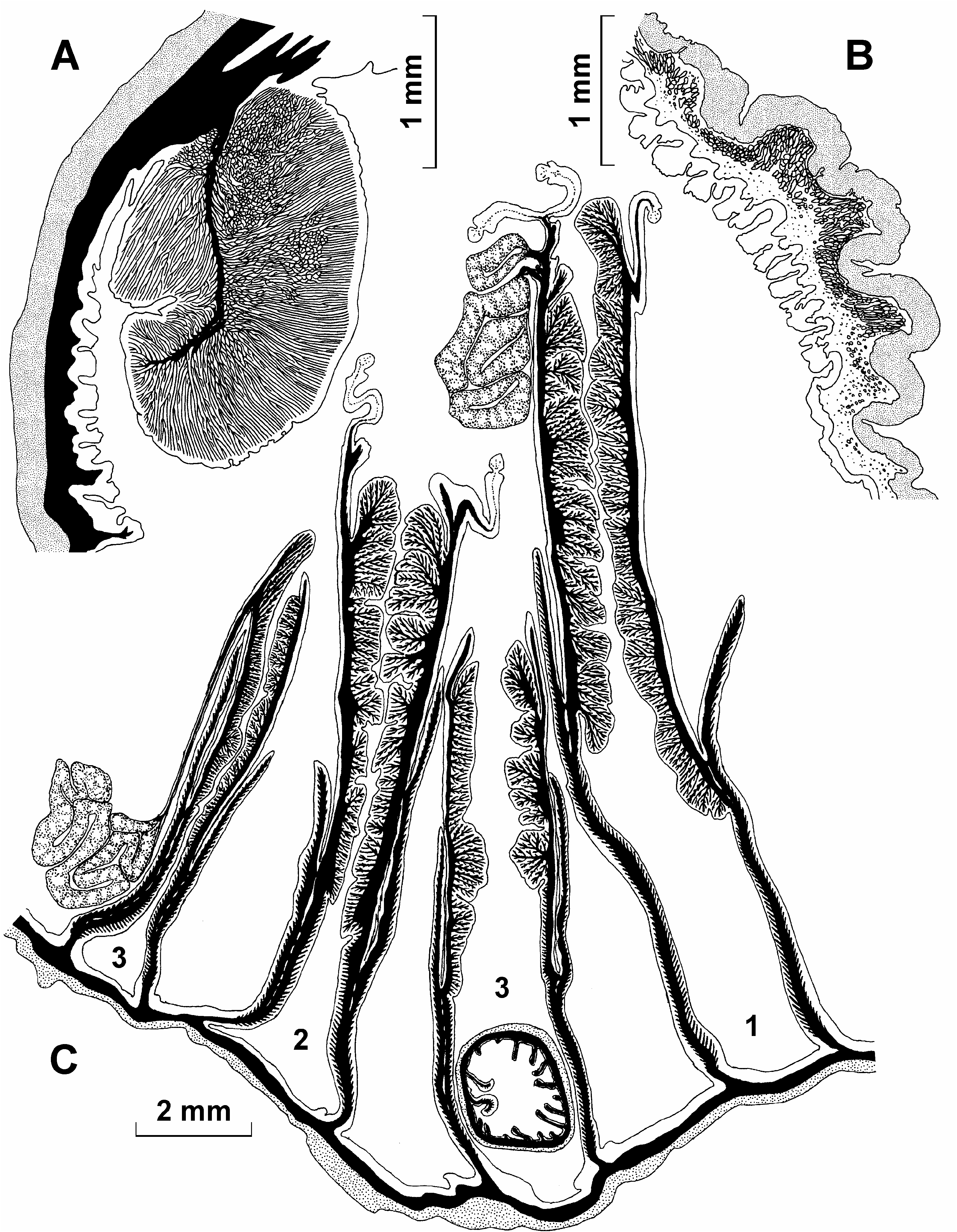
Internal structure. Circumscribed marginal endodermal sphincter is strong, circular to oval in transverse section, with one short and thin central lamella ( Figure 1A View Figure 1 ). Radial muscles of the oral disk and longitudinal muscles of the tentacles are strong, mesogloeal to ectomesogloeal ( Figure 1B View Figure 1 ). There are two deep siphonoglyphs supported by directives. Mesenteries usually are arranged decamerously and regularly, in three cycles (10+10+20 pairs), the last cycle may be incomplete. Two of the examined specimens had 9+9+18 pairs. The largest specimen had an additional pair of the fourth cycle (10+10+16+1). Some of the small mesenteries of the last cycle do not reach the margin and therefore the number of tentacles is slightly smaller than the total number of mesenteries. Mesenteries of the first, the second, and some of the third order may be perfect. The gonads in most examined specimens are poorly developed, detected mostly on mesenteries of the second and third orders, but also on the proximal part of some mesenteries of the first order. Spermatic vesicles to 0.3 mm in diameter. Retractor muscles are strong and restricted, up to reniform on the mesenteries of the youngest cycles. Strong parietobasilar muscles form a long free flap. Parietal muscles on transverse sections extend from the column wall to the retractor muscle ( Figure 1C View Figure 1 ).
Numerous large (to 9 mm long and 6 mm diameter) larvae are in one specimen (holotype) collected early in summer. Larvae have numerous tentacles, 23 being found in one expanded larva. It is interesting that this specimen has only male gonads.
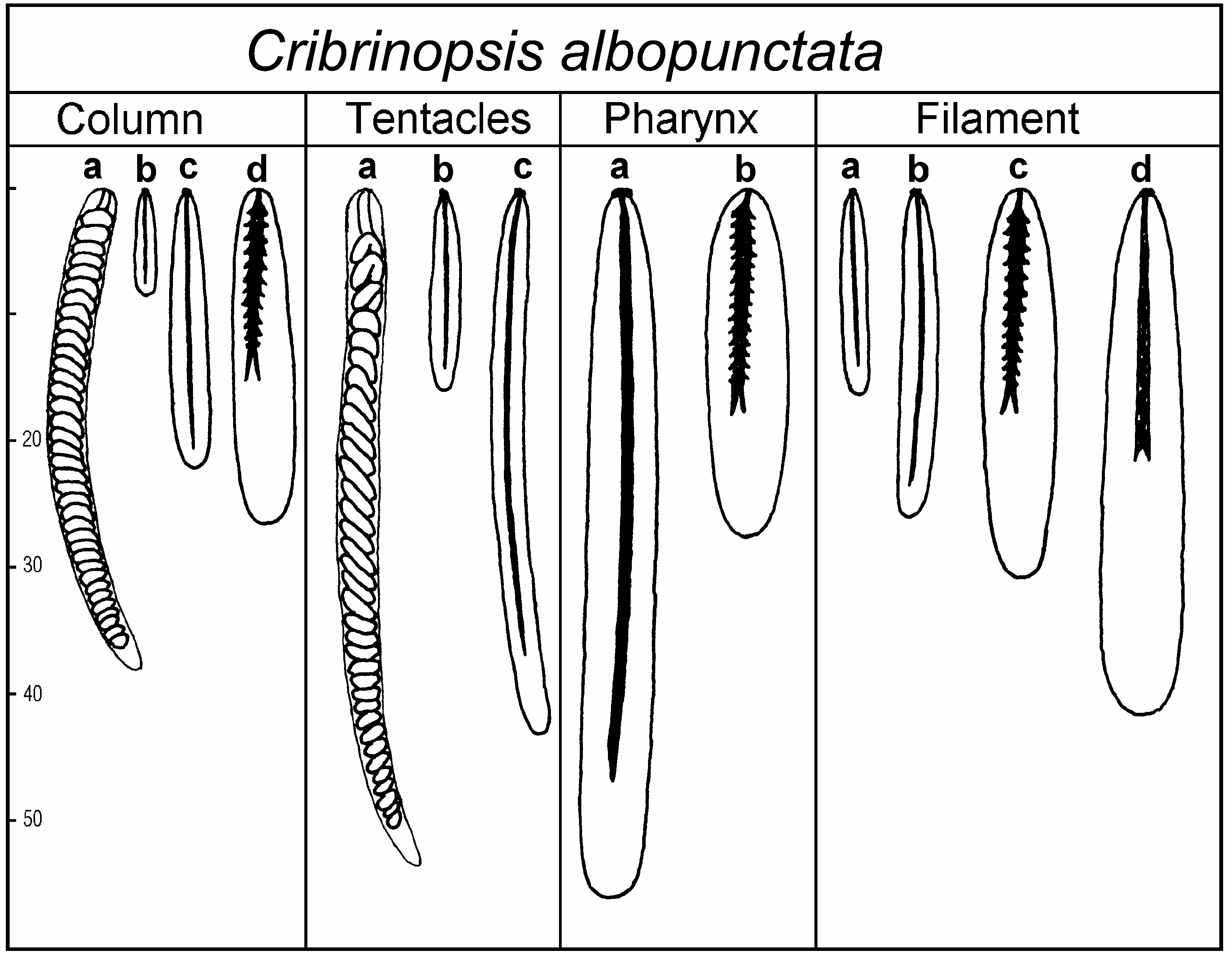
Size and distribution of cnidae (letters in parentheses refer to Figure 2 View Figure 2 , all measurements in Mm; N is the proportion of examined specimens that had a particular type of cnida; distribution of all cnidae is confirmed in sections):
Column spirocysts (a): 25–45×2.5–4 (common), N514/14 basitrichs (b): 7–16×1.5–2 (rare), N512/14 basitrichs (c): 18–30×2.5–3.5 (common), N514/14 p-mastigophores A (d): 19–30×4.5–6 (very rare), N512/14
Tentacles spirocysts (a): 22–62×2–4 (numerous), N513/13 basitrichs (b): 10–19×1.5–2.5 (rare), N55/13 basitrichs (c): 25–53×2–3 (numerous), N512/13
Actinopharynx basitrichs (a): 36–60×4–6 (numerous), N514/14 p-mastigophores A (b): 26–35×5–7 (very rare), N510/14
Filaments basitrichs (a): 11–18×1.5–2.5 (common), N511/12 basitrichs (b): 20–30×2.5–3.5 (numerous), N512/12 p-mastigophores A (c): 24–36×5–7 (common), N511/12 p-mastigophores B (d): 29–47×4.5–7 (common), N512/12
Presence of numerous spirocysts in columnar ectoderm (confirmed in sections) is one of the characteristic features of the species.
Habitat
Records from East Kamchatka are from 9 to 26 m depth and the species apparently is absent from the intertidal zone here. One specimen was found in a tide pool on the Commander Islands. Specimens always are attached to the sides or upper surfaces of stones and are never buried in sand. The column may or may not have a few attached gravel particles or broken shells.
Remarks
The present species is related to Cribrinopsis olegi sp. nov. which differs mainly in its short tentacles with spherical expanded ends, absence of spirocysts in the columnar ectoderm, and habitat ( C. olegi sp. nov. is always buried in sand). It differs distinctly from C. williamsi Carlgren, 1940 and C. fernaldi Siebert and Spaulding, 1976 which are hexamerous. Cribrinopsis similis Carlgren 1921 has a smooth column. Insufficiently described C. asiatica ( Averincev, 1967) differs in its long blunt-tipped tentacles, colour of column (yellow-white, greenish with red mosaic, brownish yellow), and colour of veruccae (brown or rose-red).
Cribrinopsis albopunctata View in CoL sp. nov. is very similar and appears to be conspecific with Pacific anemones identified by Hand (1955) as Tealia lofotensis ( Danielssen, 1890) View in CoL . However, the present specimens, as well as the other Pacific specimens recorded as T. lofotensis View in CoL (see Hand 1955; Sebens and Laakso 1977) differ distinctly from the European T. lofotensis View in CoL (see Remarks under Urticina eques View in CoL ).
The present specimens have similar sets of nematocysts, in all parts of the body studied, to those reported by Sebens and Laakso (1977) for the large specimens from Puget Sound, although the sizes of the nematocysts are slightly different and size ranges may be wider in the present specimens. The set of nematocysts reported by Hand (1955) for Californian specimens differs in more details from ours and from those of Sebens and Laakso (1977), the latter authors considered these differences to be within the expected intraspecific variation. The cnidom of the Pacific specimens is characteristic of Cribrinopsis View in CoL , but not of Urticina View in CoL . In the latter genus, unlike Cribrinopsis View in CoL , the sizes of the larger basitrichs in the tentacles and in the actinopharynx differ significantly and the size ranges do not overlap ( Figure 11 View Figure 11 ). The attribution of the present specimens to Cribrinopsis View in CoL is confirmed by occasional presence of the gonad on some mesenteries of the first cycle, although this has not been reported for the east Pacific specimens and the cnidom appears to be a more reliable feature to distinguish Cribrinopsis View in CoL from Urticina View in CoL than the distribution of gonads.
The specimens from Puget Sound are much larger than the present specimens, with the column in full extension reaching 22× 18 cm and with more tentacles, 103–135 ( Sebens and Laakso 1977). The specimens from California are a similar size to the present specimens but have more tentacles, up to 160 ( Hand 1955) and from about 94 to 154 ( Wedi and Dunn 1983). Brooding has not been observed in American specimens although large larvae were found in one of the present specimens. Despite the differences referred to above, the present specimens appear to be conspecific with the Puget Sound T. lofotensis: Sebens and Laakso, 1977 , and probably also with the Californian T. lofotensis: Hand, 1955 .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Cribrinopsis
| Sanamyan, N. P. & Sanamyan, K. E. 2006 |
Tealia lofotensis: Hand 1955 , p 80
| Sebens KP & Laakso G 1977: 162 |
| Hand C 1955: 80 |
Madoniactis lofotensis
| Danielssen DC 1890: 47 |