
Eucalliax Manning & Felder, 1991
|
publication ID |
https://doi.org/ 10.5281/zenodo.209484 |
|
DOI |
https://doi.org/10.5281/zenodo.5618128 |
|
persistent identifier |
https://treatment.plazi.org/id/D03A87F0-FFBE-FFC3-FF5E-85751D846A84 |
|
treatment provided by |
Plazi |
|
scientific name |
Eucalliax Manning & Felder, 1991 |
| status |
|
Genus Eucalliax Manning & Felder, 1991
Type species. Callianassa quadracuta Biffar, 1970 .
Remarks on taxonomy. The taxonomy of Eucalliax is rather complex. The genus was established by Manning & Felder (1991) for several species previously assigned to the genus Calliax de Saint Laurent, 1973. They noted that chelipeds are equal and similar in Eucalliax , whereas Calliax possesses unequal chelipeds. Ngoc-Ho (2003) provided a detailed account on the differences between the two genera both in soft-part and hard-part morphology. She considered chelipeds in Eucalliax subequal and similar, whereas in Calliax , chelipeds are unequal, and the minor has "fixed finger shorter than and separated from the dactylus by a wide gap, bearing a large triangular proximal tooth" ( Ngoc-Ho 2003: 490). She also erected a new genus Calliaxina Ngoc-Ho, 2003 for three species previously treated as Calliax : C. novaebritanniae ( Borradaile, 1900) , C. punica (de Saint Laurent & Manning, 1982) and C.
sakaii (de Saint Laurent & Le Loeuff, 1979). For Calliaxina , subequal, similar and laterally compressed first pereiopods are typical, whereas in Calliax , they are also lateraly compressed, but unequal and dissimilar ( Ngoc-Ho 2003). Sakai (1999, 2005) recognized Eucalliax and Calliaxina as junior synonyms of Calliax . Later, he ( Sakai 2011) recognized all of them as valid, but his concept of Eucalliax and Calliaxina differs markedly from that of Ngoc-Ho (2003). He restricted Eucalliax to Callianassa quadracuta Biffar, 1970 only, and for Calliaxina listed C. aequimana ( Baker, 1907) , C. bulimba ( Poore & Griffin, 1979) , C. jonesi ( Heard, 1989) , C. mcilhennyi ( Felder & Manning, 1994) and C. panglaoensis ( Dworschak, 2006) besides C. novaebritanniae , C. punica and C. sakaii . We follow Ngoc-Ho (2003) rather than Sakai (2011) in recognizing only three species of Calliaxina . We consider the rest of above mentioned species as members of Eucalliax , thus following works of Manning & Felder (1991), Felder & Manning (1994) and Dworschak (2005, 2006). In this respect the material presented herein with its subequal and similar chelipeds falls within the range of the genus Eucalliax . It possesses massive chelipeds which is in contrast to Calliaxina sensu Ngoc-Ho (2003) with laterally compressed chelipeds.
Remarks on the fossil record. All three Eucalliacinae genera discussed above have been identified in the fossil record. Concerning distinguishing Calliax and Eucalliax from each other in the fossil state, Schweitzer et al. (2003: 12) mentioned the nature of the lower margin of merus, which is serrated in Calliax and without serration in Eucalliax . Unfortunately, this does not seem to be consistent. For instance Biffar (1970: 42) mentioned presence of denticulation in C. quadracuta which is the type species of Eucalliax . In E. pseudorakosensis comb. nov. as recognized herein, the denticulation is also present, although only in large specimens ( Fig. 10 View FIGURE 10 E).
The fossil record of Calliax is obscure. Callianassa whiteavesi Woodward, 1896 from the Campanian of British Columbia and Alberta, Canada was reassigned to Calliax by Schweitzer et al. (2003).
Recently Feldmann et al. (2011) reported a few claw fragments as "Cheliped Form B" of indeterminate callianassoid. The material shows remarkable similarities to the genus Calliax as defined by Ngoc-Ho (2003). Interestingly, a minor chela is also preserved ( Feldmann et al. 2011: fig. 5E) exhibiting fixed finger shorter than dactylus and separated from it by a wide gap with a proximal tooth, a combination of characters which has been so far documented in Calliax only ( Ngoc-Ho 2003).
Swen et al. (2001) reported a single fragmentary right propodus from the Maastrichtian of the Netherlands classified as Calliax ? sp. It is very difficult to assign this specimen to any genus of the subfamily Eucalliacinae without evidence of both chelipeds.
Van Bakel et al. (2006) listed in a table of Cenozoic decapods from Belgium the presence of Calliax nov. sp. in the Miocene strata. The systematic placement of the material has recently been reconsidered to represent a new member of the family Axiidae ( Fraaije et al. 2011) .
Eucalliax was reported from the Eocene, Miocene and Pliocene of Japan: E. yoshihiroi Karasawa, 1992 ; E. yatsuoensis ( Karasawa, 1993) ; and E. miyazakiensis Karasawa, 1993 respectively. Kato (1996) reported several specimens classified as Eucalliax sp. from the Miocene of Japan. These specimens, however, possess tubercles on the lateral surface of propodus, which is highly unusual for the genus. Moreover, they were associated with Callianopsis spp., for which such tuberculation is quite typical. Kato (1996: 507) stated that Eucalliax sp. is distinguishable from Callianopsis spp. "by the absence of a large, triangular tooth on the proximal part of the fixed finger, and having a remarkably small fixed finger on the left chela". These characters can not be considered as important for generic assignment (see Hyžný & Schlögl 2011: 326). In fact, the presence and absence of a triangular tooth on the fixed finger may mirror sexual dimorphism in Callianopsis de Saint Laurent, 1973 as already documented by Schweitzer Hopkins & Feldmann (1997) and Hyžný & Schlögl (2011). Kato (1996) did not state why he had placed the specimens in Eucalliax .
Several occurrences of the genus have been reported also from the Eocene of Italy. Beschin et al. (2002) described Eucalliax vicetina Beschin, Busulini, De Angeli & Tessier, 2002 , based on virtually entire major and minor chelipeds. Finally Beschin et al. (2009) reported a single incomplete propodus referred to Eucalliax sp. from the Lower Eocene of Italy. Its morphology, however, strongly suggests the assignment to ctenochelid genus Ctenocheles Kishinouye, 1926 View in CoL . The presence of Ctenocheles sereaensis Beschin, De Angeli & Zorzin, 2009 in the same strata reported in the same paper may confirm the here presented opinion.
It should be noted that virtually none of the above mentioned Calliax and Eucalliax occurrences have been reexamined with respect to the work by Ngoc-Ho (2003). If following the genus concept of Eucalliacinae taxa presented by Ngoc-Ho (2003), all the fossil material assigned at one time to these genera should be revised to confirm or reject their current generic assignment.
Calliaxina has been identified in the fossil record only recently (Hyžný, in prep.). Exhaustive treatment on the distinction between Eucalliax and Calliaxina in the fossil record is beyond the scope of the paper and will be published elsewhere (Hyžný, in prep.).
Eucalliax pseudorakosensis (L ő renthey in L ő renthey & Beurlen, 1929) new combination ( Figs 7–11 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Calianassa pseudorakosensis Lőrenthey in Lőrenthey & Beurlen, 1929: 69, pl. 2 figs. 16–18.
Callianassa pseudorakosiensis . — Glaessner, 1929: 88; Schweitzer et al., 2010: 36.
Callianassa cf. pseudorakosensis . — Bachmayer, 1953: 241; Müller, 1998: 10.
Callianassa View in CoL cf. jahringensis Glaessner, 1928: 166, pl. 3 figs. 2, 2a; Glaessner, 1929: 83; Müller, 1984: 53. " Callianassa View in CoL " pseudorakosensis . — Müller, 1976: 507; Hyžný, 2011a: 41, fig. 4A, B, Table 1 View TABLE 1 , 2 View TABLE 2 .; Hyžný, 2011b: 167, Table 1 View TABLE 1 . ‘ Callianassa View in CoL ’ pseudorakosensis . — Müller, 1984: 52, pl. 3 figs. 4–5, pl. 4 figs. 1-6, pl. 5 figs. 1–4. Non Callianassa pseudorakosensis . — Radwański & Wysocka, 2004: 390, pl. 9 fig. 2.
Emended diagnosis. Heterochelous eucalliacine ghost shrimp with chelipeds without pronounced tuberculation. Merus of major cheliped ovoid with lower margin possesssing faint denticles; carpus about as long as high, squarish or rhomboidal in shape, unarmed with keeled upper and lower margins. Palm massive, subequal in length and height with keeled upper and lower margins; fixed finger with or without triangular blunt tooth; distal margin of propodus finely serrated; weak unarmed excavation extending from below the articulation with the dactylus forming a weak keel. Dactylus heavy, curved, quadrate on the cross section proximally, armed on cutting edge with blunt tooth proximally and keel forming a distal tooth. Minor cheliped smaller in size than major, dissimilar in shape, carpus longer than high, triangular in shape, upper margin keeled and arcuate; propodus higher than long, upper and lower margins distinctly keeled; fixed finger high, cutting edge slightly convex distally, dactylus slender, curved, unarmed.
Emended description. First pereiopods with major and minor cheliped strongly developed, slightly unequal in size, dissimilar in shape of propodus and dentition of fingers; major cheliped located on either right or left side of body. Two different morphotypes were identified in the nature of major propodus; morphotype with triangular tooth positioned at the base of fixed finger ( Fig. 7 View FIGURE 7 A) and morphotype without tooth ( Fig. 7 View FIGURE 7 B).
Major cheliped massive and strongly calcified; ischium slender ( Fig. 10 View FIGURE 10 C), upper margin sinuous, lower margin with row of tiny denticles; merus ovoid, longer than high (L/H = 1.8–2.0), highest at midlength, lower margin straight or slightly convex, with tiny denticles, upper margin slightly convex, smooth, lateral surface with keel positioned around the middle (e.g. Figs 10 View FIGURE 10 E, G, H); carpus about as long as high, highest distally, about as long as merus, squarish or rhomboidal in shape ( Figs. 7 View FIGURE 7 A, 10), unarmed, lower margin arcuate, upper and lower margins keeled ( Figs 8 View FIGURE 8 I, O), terminating distally in blunt corners, margins of proximo-lower corner forming almost right angle; propodus heavy, palm approximately as long as high, lateral surfaces smooth, upper and lower margins keeled, keel of lower bent slightly inward, becoming ill-defined beyond midlength and absent on fixed finger; fixed finger thick, prehensile margin unarmed (morphotype without tooth) or armed with one well separated tooth in midlength (morphotype with tooth), serrated with tiny denticles distally, otherwise unarmed, terminating in rounded tip, tip bent slightly upward; distal margin of propodus finely serrated; weak unarmed excavation extending from below the articulation with the dactylus to below the tooth on mesial face, thus forming a faint keel; dactylus heavy, curved, quadrate on the cross section proximally ( Fig. 8 View FIGURE 8 Q), armed on cutting edge with blunt tooth proximally ( Fig. 8 View FIGURE 8 J) and keel forming a distal tooth distally pointing downward near the tip of dactylus (e.g. Figs 7 View FIGURE 7 C, 8F), tip sharp and slightly hooked.
Minor cheliped smaller in size than major, dissimilar in shape, carpus longer than high, triangular in shape ( Figs 7 View FIGURE 7 D, 9E, F), upper margin keeled and arcuate; articulation between carpus and propodus more than 90º; propodus higher than long ( Fig. 9 View FIGURE 9 ), upper and lower margins distinctly keeled; fixed finger high, cutting edge slightly convex distally, the tip narrowing abruptly, pointed and bent slightly upward; dactylus slender, curved, unarmed.
Variations. There are two distinct forms, occurring on both right and left chelae, most probably due to a dimorphism. One morphotype has a triangular tooth at the base of the fixed finger, the other one has no such tooth; this dimorphism was already recognized by Lőrenthey & Beurlen (1929) and Müller (1984). A quick scan of published figures showed that at least in some taxa, such dimorphism may mirror sex. A toothed fixed finger seems to be present mostly in females, whereas a morphotype without the tooth is more typical for males as can be documented in the following examples: a distinctly toothed fixed finger has been figured in female specimens of E. aequimana ( Poore & Griffin 1979: fig. 12d), E. kensleyi Dworschak, 2005 ( Dworschak 2005: figs. 3, 4), and E.
mcilhennyi ( Felder & Manning 1994: fig. 1); a morphotype without a toothed fixed finger has been figured in male specimens of E. aequimana ( Poore & Griffin 1979: fig. 12l), E. bulimba ( Poore & Griffin 1979: figs. 21d, e), E. jonesi ( Heard 1989: figs. 1, 4b; Felder & Manning 1994: fig. 5e) and E. mcilhennyi ( Felder & Manning 1994: figs. 2b, c). On the other hand, no such pattern has been observed in E. cearaensis Rodrigues & Manning, 1992, and E. panglaoensis . Interestingly, sexual dimorphism has been observed in E. panglaoensis , although in different pattern. In this species chelipeds of females and small males are almost equal in size, whereas in large males they become slightly unequal ( Dworschak 2006: 356, fig. 7); the triangular tooth is present in major chelae in both sexes. Concerning E. cearaensis , it is questionable whether it is representative for the genus; Sakai (2011) erected for this species a new genus Eucalliaxiopsis .
We do not explain unequivocally the variability in the nature of major propodus of E. pseudorakosensis comb. nov. as a consequence of sexual dimorphism. It should, however, be mentioned that the same pattern of dimorphs have been recognized in the ctenochelid genus Callianopsis as mirroring sexual dimorphism; thus, the presence of a tooth at the base of the fixed finger is indicative of females, whereas males have no such tooth ( Schweitzer Hopkins & Feldmann 1997; Hyžný & Schlögl 2011). In this context it is worth mentioning that recent molecular analysis by Felder & Robles (2009) has shown a rather close phylogenetic relationship between Ctenochelidae Manning & Felder, 1991 View in CoL and Eucalliacinae .
Variations in the nature of the dactylus have also been observed. Usually smaller specimens have the distal tooth on the occlusal margin of dactylus rather blunt, or even not developed at all, however, in that case a swollen area in the place is present. In larger specimens the tooth becomes sharper and more distinct ( Fig. 7 View FIGURE 7 C). The same can be said about the keel on the occlusal margin of dactylus itself, which is very well pronounced in larger specimens. This type of variation can be correlated with size/age of the individual.
Material examined. FI M.20 (holotype, right major propodus, morphotype with tooth, figured in Lőrenthey & Beurlen 1929: pl. 2, fig. 16), FI M.2347 (right major propodus, morphotype with tooth); KGP-MH DH-016–KGP- MH DH-040, KGP-MH DH-040, KGP-MH DH-043, KGP-MH DH-044, KGP-MH DH-047, KGP-MH DH-051, KGP-MH DH-053, KGP-MH DH-054, KGP-MH DH-057, KGP-MH DH-059, KGP-MH DH-062, KGP-MH DH- 0 63, KGP-MH DH-069, KGP-MH DH-069; M.86.530 (collective number); SNM Z- 7943 (collective number), SNM Z- 16419 (collective number), SNM Z- 16433 (collective number), SNM Z- 21373; SNM Z- 37554– SNM Z- 37569; PAL 2011.22–PAL 2011.28; PCMH-004–PCMH-007, PCRB DH-017– PCRB DH-028. For measurements and all the details see Table 2 View TABLE 2 .
merus carpus propodus dactylus continued next page Occurrence. The species is very common in the sediments of the Middle Miocene age of the countries once forming an area of the Central Paratethys Sea. Up to now it has been reported from the late Early ('Karpatian') and Middle Miocene ('Badenian') of Hungary ( Lőrenthey & Beurlen 1929; Müller 1984), Romania ( Lőrenthey & Beurlen 1929; Müller 1984), Austria ( Glaessner 1928; Müller 1984, 1998), and Slovakia ( Hyžný 2011 a, b, this contribution).
The species often occurs in large numbers as exemplified in studied material from Dúbravská hlavica and published material from Hungarian localities Rákos and Mátraverebély-Szentkút ( Müller 1984), and from the Romanian locality Minişu de Sus (Felménes) ( Lőrenthey & Beurlen 1929; Müller 1984).
Remarks. Eucalliax pseudorakosensis comb. nov. is very easily distinguishable from all similar fossil forms. Its propodus is heavy and when viewed distally, it is very robust in its upper half (in place of articulation with dactylus), whereas the lower part is more depressed, partly also due to the arcuate and keeled lower margin curved inward. Moreover, it has a rounded cross section of the fixed finger, whereas similar Middle Miocene forms, as " Callianassa " chalmasii Brocchi, 1883 and " Callianassa " rakosiensis Lőrenthey, 1897, are oval (Lőrenthey in Lőrenthey & Beurlen 1929: 68). When dealing with more complete chelipeds, E. pseudorakosensis comb. nov. can be characterized by a squarish or rhomboidal carpus, and a dactylus armed with a distal tooth. These characters distinguish the species from all extant members of the genus, as well as from all relative fossil taxa.
Radwański & Wysocka (2004) reported from the Middle Miocene of Poland a single chela attributed to Callianassa pseudorakosensis associated with trace fossils of the ichnogenus Ophiomorpha Lundgren, 1891 . The morphology of the specimen is, however, distinctly different from E. pseudorakosensis comb. nov. The preserved propodus with articulated dactylus is somewhat fractured and slightly deformed ( Radwański & Wysocka 2004: fig.
1.2); however, the general morphology can be inferred. The upper margin seems to taper proximally unlike Eucalliax pseudorakosensis comb. nov. for which the opposite is true. The keel on the fixed finger is strongly developed; in E. pseudorakosensis comb. nov., it is developed only weakly. The dactylus is completely unlike that of E. pseudorakosensis comb. nov. in being very high and lacking the distinctive tooth positioned distally on the occlusal margin. Moreover, the specimen seems to be rather flat (laterally compressed sensu Ngoc-Ho, 2003) and not massive which is quite typical for E. pseudorakosensis comb. nov., and therefore it resembles members of the genus Calliaxina Ngoc-Ho, 2003. As the specimen has been examined only via photographs, we are hesitant to discuss its affinities further, although it is clear that it is not conspecific and possibly even not congeneric with E. pseudorakosensis comb. nov. Personal observation is needed to definitely state its systematic affinity.
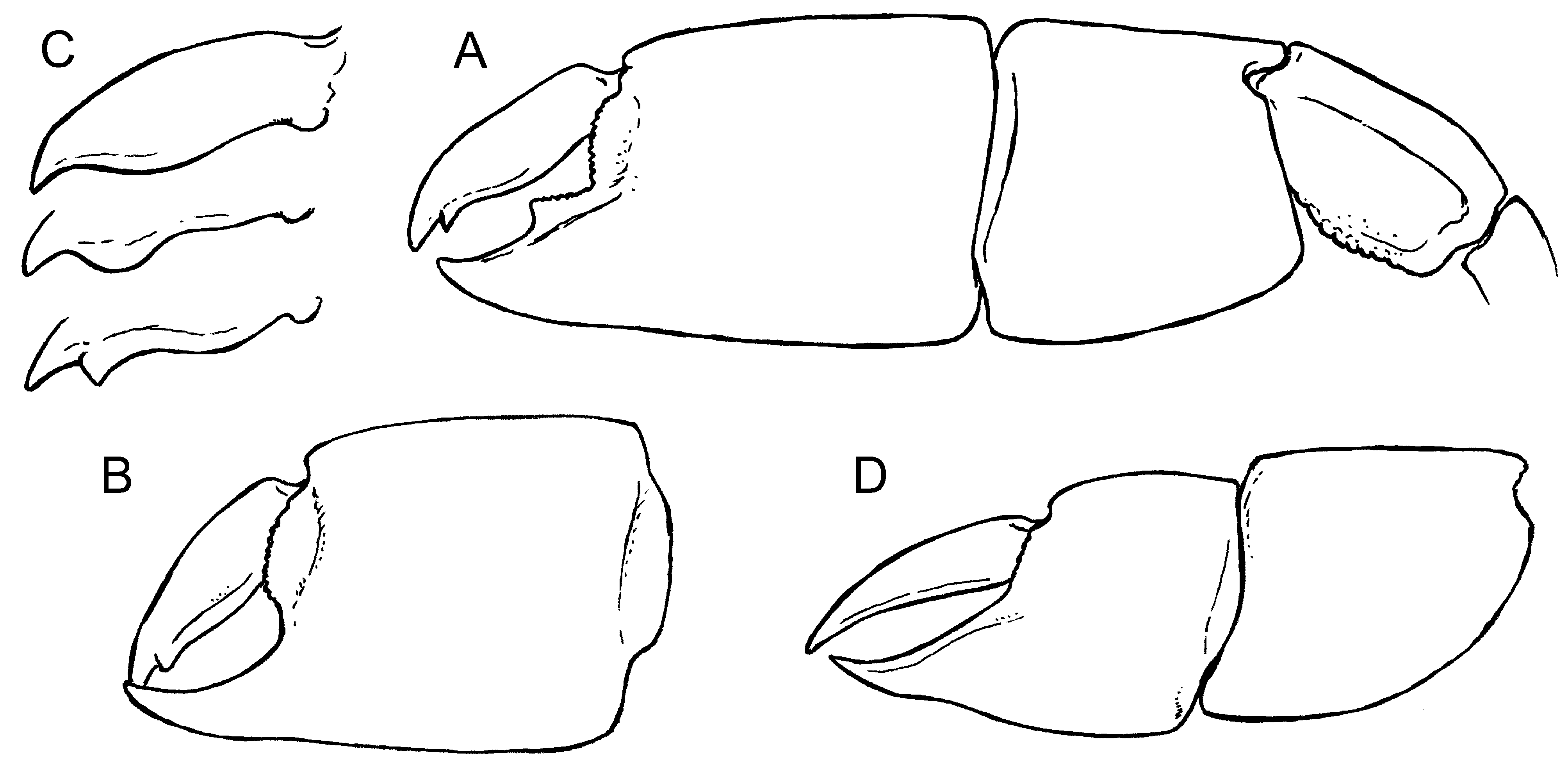

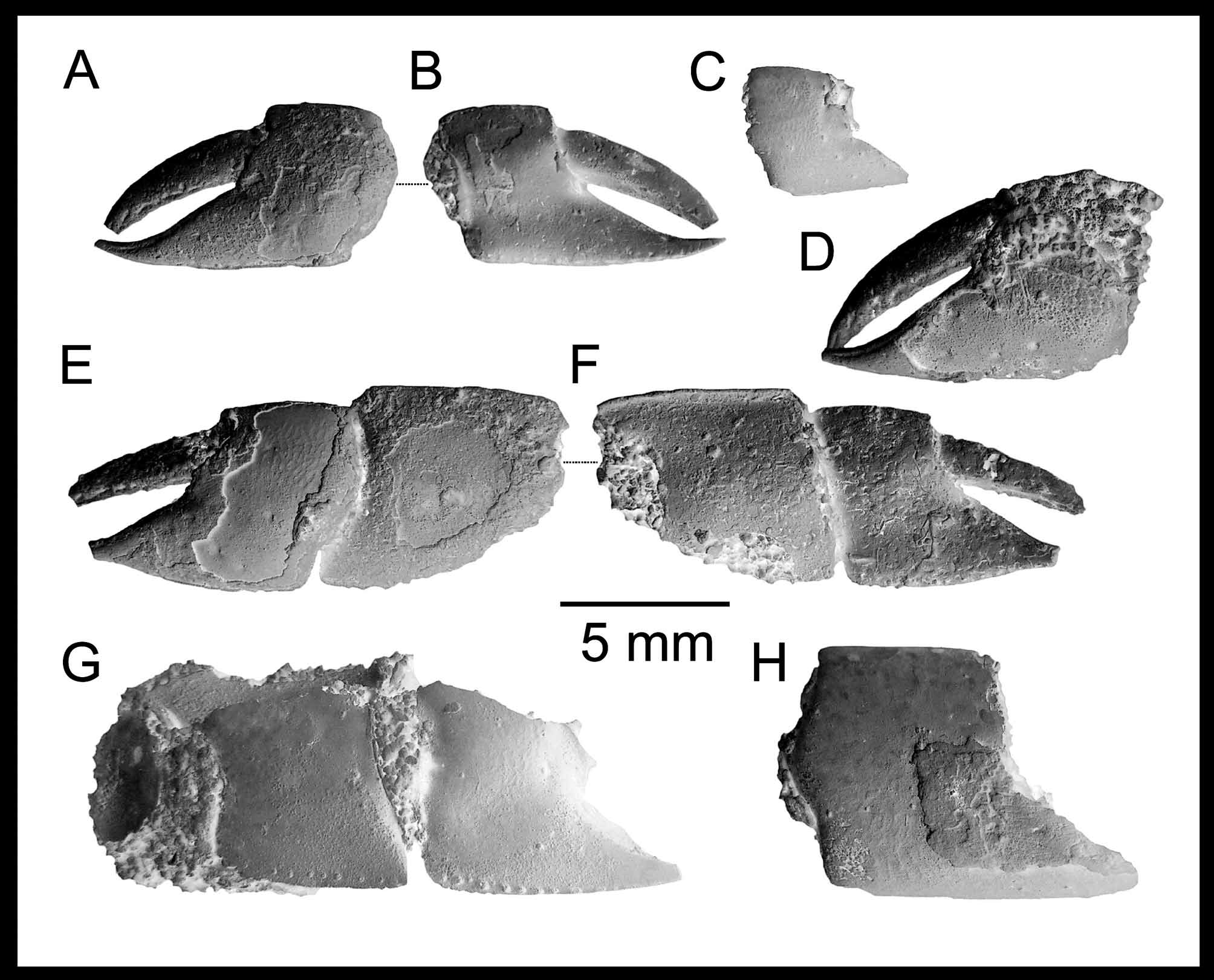


TABLE 2. Measurements of selected well preserved specimens of Eucalliax pseudorakosensis (Lőrenthey in Lőrenthey & Beurlen, 1929) comb. nov. Handedness: L = left; R = right; measurements: L = max. length; H = max. height; ~ = approximate value;> = " more than " (in case the approximation is not possible due to the state of preservation); values are in mm.
| Specimen | morphotype | handedness | L | H | L | H | L | H | L |
|---|---|---|---|---|---|---|---|---|---|
| SNM Z-21273a | with tooth | R | - | - | ~6.1 | ~6.1 | 8.0 | 8.2 | 6.7 |
| SNM Z-21273b | with tooth | R | 8.3 | 4.2 | 7.2 | 8.4 | 8.0 | 8.8 | 6.4 |
| SNM Z-21273c | with tooth | R | - | - | - | - | 8.7 | 8.4 | 6.8 |
| SNM Z-37554 | without tooth | L | - | - | - | - | 7.1 | 8.2 | - |
| SNM Z-37555 | without tooth | L | - | - | - | - | 8.2 | 7.9 | 8.0 |
| SNM Z-37556 | with tooth | R | - | - | - | - | 9.0 | 8.3 | 6.0 |
| SNM Z-37558 | with tooth | R | - | - | - | - | 11.2 | 9.0 | - |
| SNM Z-37559 | with tooth | L | - | - | - | - | 7.6 | 7.8 | ~5.0 |
| SNM Z-37560 | with tooth | L | - | - | - | - | 6.0 | 5.4 | >3.3 |
| SNM Z-37561 | minor | R | - | - | - | - | 6.0 | 8.1 | - |
| SNM Z-37562 | without tooth | R | - | - | - | - | 8.5 | 8.6 | - |
| SNM Z-37563 | without tooth | R | - | - | >7 | 9.5 | 10.7 | 10.0 | - |
| SNM Z-37564 | without tooth | L | 7.1 | - | 7.7 | 8.5 | 7.6 | 9.2 | - |
| SNM Z-37565 | without tooth | L | - | - | 7.3 | 9.1 | 9.0 | 9.4 | - |
| SNM Z-37566 | with tooth | L | - | - | - | 9.5 | - | 10.1 | - |
| SNM Z-37567 | unknown | R | - | - | 5.0 | 7.5 | 7.5 | - | - |
| SNM Z-37568 | with tooth | R | - | - | >3.8 | 5.7 | 5.4 | 5.3 | 5.3 |
| SNM Z-37569 | without tooth | R | 7.0 | 4.4 | - | - | 9.0 | >8.0 | >8.4 |
| KGP-MH DH-016 | without tooth | L | - | - | - | - | >8.0 | 10.1 | - |
| KGP-MH DH-018 | without tooth | R | - | - | - | - | 11.1 | 10.0 | - |
| KGP-MH DH-019 | without tooth | R | - | - | - | - | 10.7 | 10.2 | - |
| KGP-MH DH-020 | unknown | R | - | - | - | - | 8.4 | 7.7 | - |
| KGP-MH DH-021 | with tooth | L | - | - | - | - | >7.5 | 7.5 | - |
| KGP-MH DH-022 | with tooth | L | - | - | - | - | >10.5 | 9.4 | - |
| KGP-MH DH-023 | with tooth | L | - | - | - | - | 9.0 | 8.8 | - |
| KGP-MH DH-024 | with tooth | L | - | - | - | - | 5.7 | 8.2 | - |
| KGP-MH DH-025 | with tooth | L | - | - | - | - | 8.2 | 7.5 | - |
| KGP-MH DH-026 | with tooth | L | - | - | - | - | 6.2 | 7.3 | - |
| KGP-MH DH-027 | with tooth | R | - | - | - | - | 10.4 | 10.1 | 9.2 |
| KGP-MH DH-028 | without tooth | R | - | - | - | - | 9.5 | 10.3 | - |
| KGP-MH DH-029 | without tooth | R | - | - | - | - | 7.0 | 7.2 | - |
| KGP-MH DH-030 | with tooth | R | - | - | - | - | >9.0 | >7.5 | - |
| KGP-MH DH-033 | with tooth | L | - | - | - | - | 7.1 | 7.1 | 6.0 |
| KGP-MH DH-034 | unknown | R | 6.2 | 5.2 | 5.5 | 7.2 | ~6.0 | 7.3 | - |
| KGP-MH DH-036 | unknown | R | - | - | 8.2 | 8.7 | - | - | - |
| KGP-MH DH-037 | unknown | R | - | - | 8.0 | 10.0 | - | - | - |
| KGP-MH DH-039 | with tooth | R | 9.5 | 5.1 | 9.6 | 9.4 | 10.0 | 9.3 | ~8.0 |
| KGP-MH DH-040 | without tooth | R | - | - | - | - | 10.9 | 10.0 | >7.5 |
| KGP-MH DH-043 | without tooth | L | - | - | - | - | 10.7 | 9.5 | - |
| KGP-MH DH-051 | with tooth | L | - | - | - | - | 8.2 | 8.0 | - |
| SNM |
Slovak National Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Eucalliax Manning & Felder, 1991
| Hyžný, Matúš & Hudáčková, Natália 2012 |
Callianassa cf. pseudorakosensis
| Muller 1998: 10 |
| Bachmayer 1953: 241 |
Calianassa pseudorakosensis Lőrenthey
| Lorenthey 1929: 69 |
Callianassa pseudorakosiensis
| Schweitzer 2010: 36 |
| Glaessner 1929: 88 |
Callianassa
| Hyzny 2011: 41 |
| Hyzny 2011: 167 |
| Radwanski 2004: 390 |
| Muller 1984: 53 |
| Muller 1984: 52 |
| Muller 1976: 507 |
| Glaessner 1929: 83 |
| Glaessner 1928: 166 |