
Eulimnadia pampa, Marinone, María Cristina, Urcola, Juan Ignacio & Rabet, Nicolas, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4158.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:A4E2FE08-EF9F-47DB-B542-ABB2697BBB80 |
|
DOI |
https://doi.org/10.5281/zenodo.6080936 |
|
persistent identifier |
https://treatment.plazi.org/id/D03B7850-FFB6-FFFD-FF0F-7DC43A75627E |
|
treatment provided by |
Plazi |
|
scientific name |
Eulimnadia pampa |
| status |
sp. nov. |
Eulimnadia pampa View in CoL sp. nov.
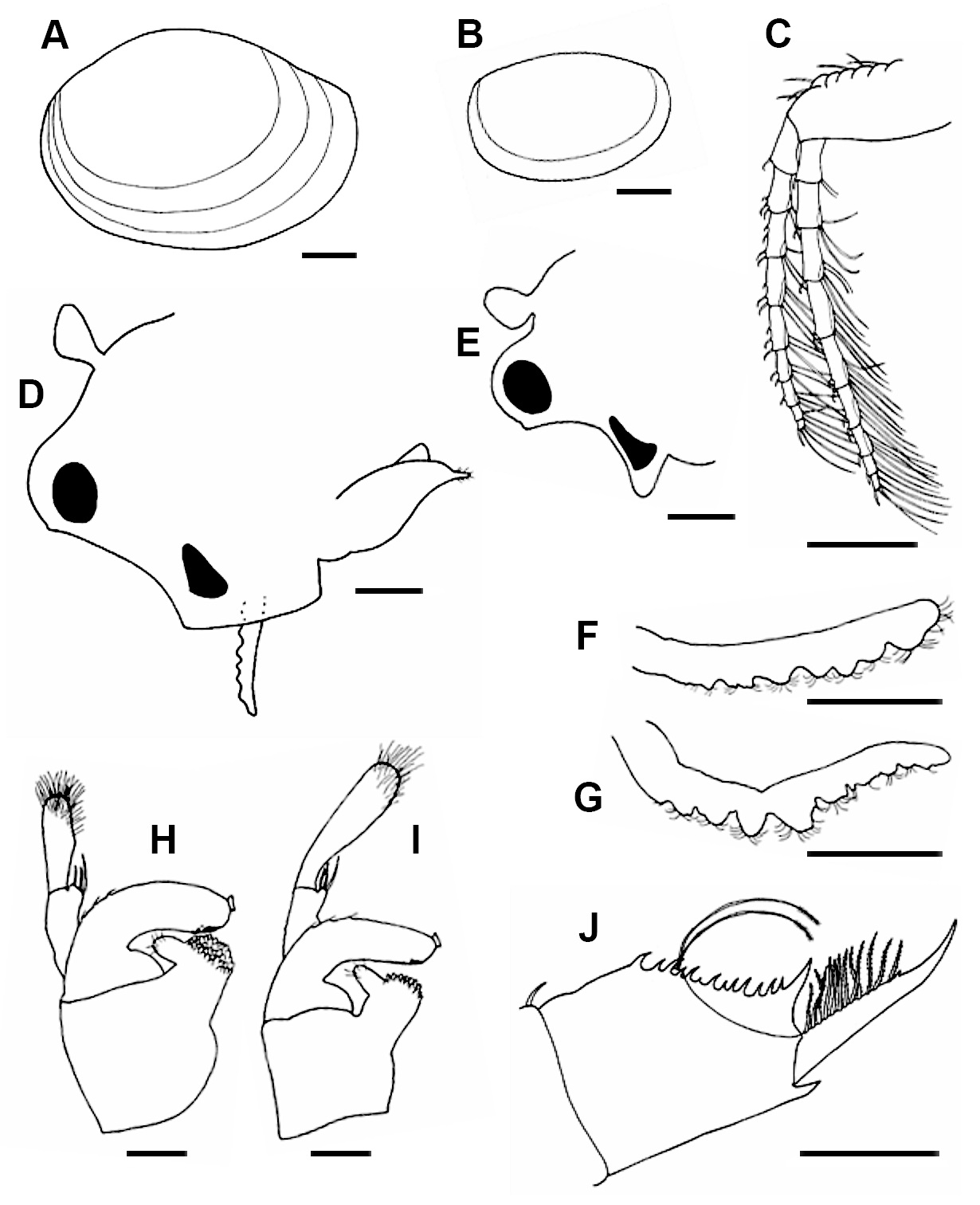
( Figs. 1 View FIGURE 1 , 4 View FIGURE 4 A–D)
Etymology. The specific epithet “ pampa ” is a noun in apposition that refers to the eco-region where the new species was found.
Type locality. Buenos Aires City ( CABA): rain pools at 34°33’49.47" S, 58°25’18.36" W, 12 m asl. Rain pools on clayey soil covered by grass, partially shaded by an artificial forest of Eucalyptus sp. trees. GoogleMaps
Type material. Holotype: ovigerous putative hermaphrodite preserved in 80% ethanol, 15-II-2001, CABA, Paseo de las Américas Park, coll.: M.S. Fontanarrosa (MACN-In 39872).
Allotype: male, same date, locality and collector as holotype (MACN-In 39873).
Paratypes: 7 hermaphrodites (2 ovigerous) and 4 males, Buenos Aires Province, Ezeiza , 04-II-2015, coll.: J.I. Urcola (MACN-In 39874). Two ovigerous hermaphrodites and one male, CABA, De la Raza (or Bosque Alegre) Park , 28-X-1998, coll.: M.C. Marinone (MACN-In 39875). One male, CABA, República del Ecuador Park, 30-I- 2001, coll.: M.C. Marinone (MACN-In 39876). Two ovigerous hermaphrodites and one male, CABA, Paseo de las Américas Park , 06-II-2001, coll.: M.C. Michat (MACN-In 39877). One ovigerous hermaphrodite, CABA, Paseo de las Américas Park , 15-II-2001, coll.: P.L.M. Torres (MACN-In 39878). One ovigerous hermaphrodite, CABA, República del Ecuador Park, 07-III-2 0 0 1, coll.: M.S. Fontanarrosa (MACN-In 39879). One male, CABA, Paseo de las Américas Park, 22-III-2002, coll.: P.L.M. Torres (MACN-In 39880) . 18 eggs mounted for SEM, CABA, Tres de Febrero Park, 11-XII-2003, coll.: R. Suárez (MACN-In 39881). Additional paratypes were deposited in the MNHN, France (14 ovigerous hermaphrodites, 3 males, MNHN-IU-2014-12862) and in the Invertebrates Collection of the Departamento de Biodiversidad y Biología Experimental, FCEyN, UBA (IC-DBBE-UBA).
Material examined. Argentina: Buenos Aires City ( CABA): rain pools in urban parks, at 34°33'49.47" S, 58°25'18.36" W, 12 m asl, coll.: M.C. Marinone, M. C. Michat, P. L.M. Torres, and M.S. Fontanarrosa, 28-X-1998, 18-XII-1998, 30-I-2001, 06-II-2001, 14-II-2001, 15-II-2 0 0 1, 06-III-2001, 07-III-2001, 27-III-2001, and 28-III- 2001, n = 263; vegetated rain pool in the former “ Bañado de Flores ”, at GoogleMaps -34°40'00" S, 58°27'25" W, 3 m asl, coll: A.O. Bachmann, 08-I-1956, n = 28 (poorly preserved), IC-DBBE-UBA. Buenos Aires Province: Partido de Ezeiza : rain pools at 34°46'05.14" S, 58°32'53.84" W, 19 m asl, collector: J.I. Urcola, 23-IX-2013, 03-X-2013, 07-XI-2013, 28-XI-2013, 10-I-2014, 23-I-2014, 08-II-2014, 06-III-2 0 14, 20-III-2014, 18-I-2015, and 26-XII-15, n = 118; Partido de Bragado: rain pool in Estancia La Escondida, at 35°14'31.86" S, 60°42'47.81" W, 61 m asl, coll.: M.C. Michat and P.L.M. Torres, 18-I-2006, n = 1 GoogleMaps . Entre Ríos Province: Colón Department: temporary pool by Carballo stream, at 32°06'13.94" S, 58°14'40.92" W, coll.: M.C. Marinone, 21-XII-2001, n = 14. GoogleMaps
Diagnosis. Typical Eulimnadia as defined by Rogers et al. (2012). Eggs subspherical, ornamented by deep elongate hexagonal flat bottom depressions, separated by sharp ridges with irregularly serrated margins and spiniform projections.
Description. Putative hermaphrodite. Carapace compressed, unevenly ovoid, lacking umbone; transparent to light amber; integument soft covered by minute punctations; dorsal hinged margin markedly domed in larger individuals, higher than in males, highest at about 1/3 of the total length of the carapace, angulated both anteriorly and specially posteriorly; 1 to 9 (plus margin) well defined growth lines, more closely spaced anteriorly than posteriorly ( Fig. 1 View FIGURE 1 A).
Head. Rostrum angular, obtuse, lower angle ending into a pointed apex, shallow sinus between the ocular tubercle (OT) and the rostrum apex; naupliar eye large elongate triangular, slightly smaller than the eye; compound eye large, occupying most of ocular tubercle, which is much larger than frontal organ (FO) (OT/FO = 1.88); small notch in ocular tubercle frontal region indicating position of frontal pore; frontal organ with apical depression, pedunculate, subequal in size to compound eye (CE) (CE/FO = 0.96); labrum acuminate, ending in slender cylindrical section ( Fig. 1 View FIGURE 1 D).
First antenna unannulated, pseudoarticulated in middle part, bearing about seven frustoconical lobes of different size in a longitudinal row, three basally and four distally, plus a terminal sensory setae cluster, all lobes bearing numerous curved sensory setae or aesthetascs ( Fig. 1 View FIGURE 1 F). Length: 431–670 µm, width: 67–105 µm, relative length: extending to the base of second antennae.
Second antenna. Exopodite (anterior flagellum, EX) and endopodite (posterior flagellum, EN) with 9 flagellomeres each (separation of the distal flagellomeres often incomplete), EN length/EX length = 1.15. Each flagellomere armed with variable number of spines (up to 4) dorsally and a variable number of long plumose setae and an elongate keel ventrally ( Fig. 1 View FIGURE 1 C).
Thorax (trunk) provided with 16 to 18 thoracopod-bearing segments; first pair of thoracopods with endite V subequal in length to endopodite; exopodite proximal branch slightly curved and half the length of the distal branch ( Fig. 2 View FIGURE 2 A); 8th and 9th or 9th and 10th or 10th and 11th pairs of thoracopods bearing elongated egg carrying epipodite, about four times longer than the endopodite ( Fig. 2 View FIGURE 2 B); last thoracopod pairs reduced; 7 to 12 posterior segments bearing dorsal tufts of a few stiff setae.
Telson subpentagonal, with dorsal and ventral margins subparallel; ventroposterior angle bearing a posteriorly directed spiniform projection at cercopod base (CP); cercopods a little longer than telson ventral margin; posterior spine rows (formerly called dorsal spines) bearing 11 to 20 straight to slightly curved spines of irregular size (some are small), confluent dorsally, distal one noticeably longer and wider basally than others ( Fig. 1 View FIGURE 1 J); telsonic filaments (TF) distally plumose, shorter than cercopods (TF/CP = 0.7 in large individuals), borne from a projecting mound located between the third and fourth proximal spines of the posterior spine rows; cercopods proximal two thirds straight, distal third arcuate, bearing a dorsal row of 10 to 12 uniformly long, distally plumose setae, terminating distally with a minute spine at two-thirds of the length ( Fig. 1 View FIGURE 1 J); subapical cirrus formed by 10 to 14 irregular spinulae ( Fig. 2 View FIGURE 2 C).
Spermatophores. Dimensions: 309–350 x 159–230 µm (n = 5). Flat, amorphous (subquadrangular, subtrapezoidal or subtriangular), and irregularly lobed ( Fig. 3 View FIGURE 3 ). The pair of structures differs in shape on each side ( Fig. 3 View FIGURE 3 A, B). The smooth surfaces show no trace of egg ornamentation but have clear imprints of the endites and setae of the limbs where they were affixed ( Figs. 3 View FIGURE 3 A, B), whereas the outer surface appears smooth ( Fig. 3 View FIGURE 3 C).
Eggs. Egg diameter (including projections): 172–216 µm (n=15). Subspherical; sculptured with deep elongate polygonal (mainly hexagonal) flat bottomed depressions, separated by sharp ridges with irregularly serrated margins; spiniform projections at ridge intersections remarkably variable in length ( Figs. 4 View FIGURE 4 A–D); egg internal cross section circular ( Fig. 5 View FIGURE 5 A), cortical layer textured with minute pores ( Fig. 5 View FIGURE 5 C), internal alveolar layer of thin trabeculae (0.1–0.4 µm) separating polygonal or spherical vesicles of varying diameter, smaller (0.4–1.1 µm) near the chorion basal zone and distally larger (2.3–4.2 µm) towards the central part of the ridges ( Figs. 5 View FIGURE 5 B, C). Although external ornamentation varies among populations, even within the same area, the internal shell structure is identical ( Figs. 5 View FIGURE 5 B, C).
Male. Carapace. Gently domed dorsal hinged margin, much lower than in putative hermaphrodites, highest at a distance of 0.4 from the anterior end, slightly concave both anteriorly and posteriorly; number of concentric growth lines from 2 to 6 (including margin), closer to each other anteriorly ( Fig. 1 View FIGURE 1 B).
Head. Rostrum produced into a triangular apex (mostly occupied by a large naupliar eye), with ventral angle ~90˚; ocular tubercle and rostrum apex separated by deep sinus; ocular tubercle larger than frontal organ (OT/FO = 1.52); frontal organ with an apical depression, pedunculate, subequal is size to compound eye (CE/FO = 1.33); other characters similar to putative hermaphrodite ( Fig. 1 View FIGURE 1 E).
First antenna larger than in hermaphrodite; pseudoarticulated in its middle part, between two largest lobes, bearing about nine frustoconical lobes, four basally (proximal lobes shorter) and five distally, all lobes bearing numerous curved aesthetascs ( Fig. 1 View FIGURE 1 G). Length: 651–756 µm, width: 96–163 µm, relative length: extending to half of the length of antennomere 2 of second antennae.
Second antenna. Exopodite with about 8 antennomeres, endopodite with 9 antennomeres; EN length/EX length = 1.20; other characters as in putative hermaphrodite.
Thorax (trunk) provided with 16 thoracopod-bearing segments; last thoracopod pairs reduced; first two pairs modified as claspers (nomenclature follows Kaji et al. 2014 and Olesen et al. 1996). First clasper ( Fig. 1 View FIGURE 1 H) “large palp” (endite V) proximal palpomere curved, distal palpomere clavate, scarcely longer than palm (endite IV + corm of endite V); second clasper ( Fig. 1 View FIGURE 1 I) “large palp” proximal palpomere straight, distal palpomere clavate, about twice as long as palm. Both palps with proximal palpomere with a distal setal cluster, and distal palpomere bearing distal sensory setae; palm subquadrangular (as long as wide), lacking setae, apical club (endite IV) frustoconical, with a distal gripping area provided with numerous minute subtriangular scaliform denticles ( Fig. 2 View FIGURE 2 D) and a lateral bunch of plumose setae; movable finger (endopodite) with an expanded base, arcuate, with three dorsal spines, and a ventral preapical field provided with many rounded pits and a small apical suctorial organ ( Fig. 2 View FIGURE 2 D); “small palp” (endite IV outgrowth) arising laterally and perpendicularly from the apical club, bearing a group of terminal setae.
Telson as in putative hermaphrodite, except for fewer spines (8 to 16) in posterior spine ridges; cercopod length about 1.7 times length of telson ventral margin; cercopod with 9 to 10 long plumose setae, subapical cirrus formed by around 8 irregular spinules.
Size. Ovigerous specimens: carapace length 3.0– 6.6 mm, height 1.9–4.9 mm, height/length 0.61–0.74 (n = 103), width 1.2–2.1 mm (n = 48). Males: carapace length 2.8–5.2 mm, height (dorsal to ventral) 1.8–3.3 mm, height/length 0.6–0.7 (n = 23), width (right to left side) 0.90–1.6 mm (n = 13).
Breeding system. Based on 20 samples collected on different dates (n = 106 adults), we obtained a mean ratio of 0.21 male:female, from which we infer that E. pampa sp. nov. is androdioecious, and that the egg-carrying individuals should be considered hermaphrodites according to the criteria by Sassaman (1995) and Weeks et al. (2005, 2008).
Ecology. The new species occurs in seasonally astatic pools (30–190 m 2 in surface area, up to 40 cm deep). Associated fauna include amphibian larvae, cladocerans, copepods, ostracods, and a great diversity of aquatic insects ( Fischer et al. 2000). Under natural conditions, at mean temperatures of 18 to 24°C, juveniles appear 2 days after innundation and mature individuals after 6 days.
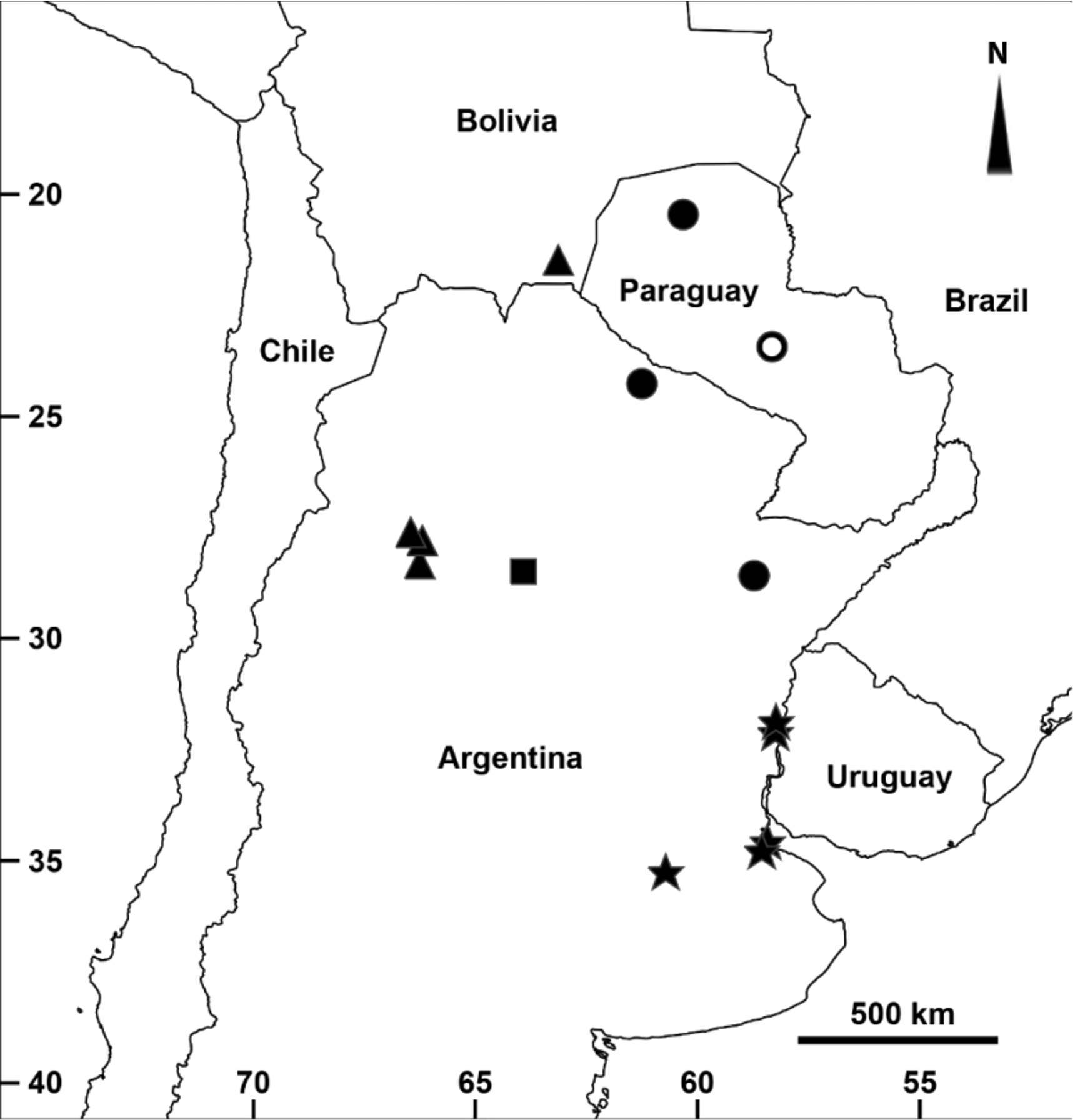
Distribution. Argentina: Buenos Aires and Entre Ríos provinces ( Fig. 6 View FIGURE 6 ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |