
Propalaeotherium gaudryi ( Lemoine, 1878 )
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2023v45a9 |
|
publication LSID |
urn:lsid:zoobank.org:pub:1C430978-5EE6-49AE-AF7C-23C710161CB7 |
|
DOI |
https://doi.org/10.5281/zenodo.8037904 |
|
persistent identifier |
https://treatment.plazi.org/id/D066B24B-5209-B676-FF20-F973DFEFF87F |
|
treatment provided by |
Felipe |
|
scientific name |
Propalaeotherium gaudryi ( Lemoine, 1878 ) |
| status |
|
Propalaeotherium gaudryi ( Lemoine, 1878)
( Figs 8 View FIG ; 9 View FIG )
Pachynolophus gaudryi Lemoine, 1878: 19 .
Pachynolophus (Propachynolophus) gaudryi – Lemoine 1891: 285- 286, figs 109-115.
Hyracotheryus [sic] – Lemoine 1891: 266, pl. XI, fig. 121. Propachynolophus gaudryi – Teilhard de Chardin 1921: 71-73, fig. 35. — Savage et al. 1965: 15-27, figs 7-8, 10-11.
Propalaeotherium gaudryi – Remy 2017: figs 1A-C, 2A-D, 10A, B, 12A-C.
LECTOTYPE ( Savage et al. 1965). — MNHN-AL5210 , fragmentary left mandible with m1-m3 .
MATERIAL. — Ageian: Maxillary (composite) with P2-M3 (L: MNHN-AL 6540); dP3 (R: MNHN-AL 5219, AL6560); P3 (R: MNHN-AL 6539); M2 (R: MNHN-AL6549; L: MNHN-AL 6533, AL6553); M1/2 (R: MNHN-AL6534, AL6536; L: MNHN-AL 6565); M3 (R: MNHN-AL6532, AL6547; L: MNHN-AL 6566); symphysis ( MNHN-AL 5214); mandible p1-p4 (L: MNHN-AL 5212); mandible p3-m3 ( MNHN-AL 5211); mandible m2-m3 (L: MNHN-AL 5213); p1 (L: MNHN-AL 5218); p3 (R: MNHN-AL6542; L: MNHN-AL 6541); p4 (R: MNHN-AL 6544); astragalus (R: MNHN. unumbered), calcaneus (L: MNHN.unumbered) .
Chavot: M1/2 (R: MNHN-CHO15569; L: MNHN-CHO 15519, CHO 15521); mandible dp1-m3 (L: MNHN-CHO14953 [cast]); mandible dp3-m2 (L: MNHN-CHO 14801); mandible p2-m3 (L: MNHN-CHO 14884); mandible p4-m3 (L: MNHN-CHO 14911); mandible m2-m3 (L: MNHN-CHO 14789); symphysis ( MNHN-CHO 14787) .
Cuis: maxillary P2-M3 (L: FSL-2096); maxillary P3-P4 (R: MNHN-CUI15540 ); P3 (R: MNHN-CUI15548 ); M1/2 (R: MNHN-CUI15537, CUI15538, CUI15546, CUI-5-L; L: MNHN-2003-2-78, MNHN-CUI15506 , CUI15513 ); dp4 (R: MNHN. 2003- 2-20); p4 (L: MNHN-CUI15510 ); m1 (R: MNHN-CUI15511; L: MNHN-CUI15504 ); m2 (L: MNHN-CUI15505 ); m1/2 (R: MNHN-CUI15545 ); m3 (R: MNHN-CUI192, MNHN- 2003- 2-18); humerus (L: MNHN.2003-2) .
Grauves: Maxillary M2-M3 (R: MNHN-GR61-L; L: MNHN-GR62-L); M1 (R: MNHN-GR60-L; L: MNHN-GR42-L, MNHN-GR46-L); M1/2 (R: MNHN-GR29-L); mandible m1-m3 (L: MNHN.unnumbered); upper canine (MNHN-GR280-L); dP3 (R: MNHN-GR7888, GR187); dP4 (L: MNHN-GR8-L); P2 (R: MNHN-GR7400, GR7401, GR276-L); upper premolar (R: MNHN-GR14-L); P3 (R: MNHN-GR7380, GR24-L, GR28-L, GR32-L; L: MNHN-GR7576, GR13-L, GR39-L); P4 (R: MNHN-GR10850, GR10-L, GR36-L, GR268-L; L: MNHN-GR7573, GR31-L, GR40- L); M1/2 (R: MNHN-GR7409, GR10847, GR10841, GR10786, GR10787, GR271-L, GR193-L; L: MNHN-GR7408, GR10839, GR10849, GR15-L); M3 (L: MNHN-GR59-L); mandible m1-m3 (R: MNHN-GR10788); dp3 (R: MNHN-GR7890; L: MNHN-GR10784); dp4 (R: MNHN-GR180); p1 (MNHN-GR7546); p2 (MNHN-GR7-L); p3 (R: MNHN-GR4-L; L: MNHN-GR48-L); p4 (R: MNHN-GR7572, GR266-L; L: MNHN-GR2-L, GR18-L); m1 (L: MNHN-GR7381); m1/2 (R: MNHN-GR20-L, GR57-L; L: MNHN-GR7571, GR26-L, GR27-L, GR55-L, GR272-L); m3 (R: MNHN-GR10776, GR34-L, GR50-L, GR52-L, GR56-L; L: MNHN-GR17-L, GR45-L, GR47-L, GR270-L); metacarpal II (R: MNHN.F.GR18004); astragalus (R: MNHN-GR10911); metatarsal III (L: MNHN.F.GR18002, MNHN.F.GR18003); metatarsal IV (R: MNHN.F.GR18001; L: MNHN.F.GR18000) .
Mancy: Maxillary P1-M3 (L: MNHN-MA14948); maxillary M1-M2 (L: MNHN-MA14795); maxillary M1-M3 (R: MNHN-MA14948); maxillary M2-M3 (L: MNHN-MA14788, MA14802); P2 (L: MNHN-MA61-L); M1/2 (R: MNHN-MA14988, MA14995; L: MNHN-MA14989, MA14990, MA14993, MA14994, MA14996, MA39-L, MA50-L, MA54-L, MA56- L); M2 (R: MNHN-MA15571); M3 (L: MNHN-MA14985); mandible dp4-m1 (R: MNHN-MA3-L); mandible p1-m3 (L: MNHN-MA9-L); mandible p3-m3 (R: MNHN-MA14902; L: MNHN-MA14907); mandible p4 (R: MNHN-MA14977); mandible p4-m3 (R: MNHN-MA14886; L: MNHN-MA10-L); mandible m1 (L: MNHN-MA14796); mandible m1-m3 (R: MNHN-MA14883 [cast]); mandible m2 (L: MNHN-MA14803); mandible m2-m3 (R: MNHN-MA14791; L: MNHN-MA14790); mandible m3 (R: MNHN-MA14797, MA14793, MA70-L; L: MNHN-MA14978); symphysis (MNHN-MA14908, MA14786); P1 (R: MNHN-MA78-L); P2 (L: MNHN-MA14842); M3 (R: MNHN-MA75-L); dp4 (R: MNHN-MA14755); p1 (L: MNHN-MA29-L); p2 (L: MNHN-MA15501); p3 (R: MNHN-MA23-L; L: MNHN-MA14739, MA15502); p4 (R: MNHN-MA27-L; L: MNHN-MA15573, MA25-L); m1 (L: MNHN-MA14986, MA14999); m1/2 (R: MNHN-MA14754, MA17-L, MA18-L, GR30-L); m3 (L: MNHN-MA21-L, MA22-L), metacarpal II (R: MNHN.F.MA18001); metatarsal III (R: MNHN.F.MA18000); metatarsal IV (R: MNHN.F.MA18002) .
Mont Bernon: p3 (R: MNHN-B5-L); p4 (R: MNHN-B10-L); m3 (L: MNHN-B4-L); metatarsal III (L: MNHN.F.MTB18000) .
Monthelon: Maxillary DP3-DP4 (L: MNHN-MTH53-L); maxillary M1-M3 (L: MNHN-MTH48-L); incisor (MNHN-MT23-L); upper canine (R: MNHN-MTH45-L; L: MNHN-MTH57-L); P4 (R: MNHN-MTH24-L); M1/2 (L: MNHN-MTH14-L, MTH44-L, MTH56-L, MTH57-L); mandible dp3-m3 (R: MNHN-MTH5-L); mandible p3-m3 (R: MNHN-MTH2-L); p4 (L: MNHN-MTH22-L), m1/2 (R: MNHN-MTH54-L); m3 (R: MNHN-MTH27-L), metatarsal II (L: MNHN.F.MTH18000) .
Saint-Agnan: Fragmentary m3 (R: MNHN-STA605-L) .
TYPE LOCALITY. — Environs d’Epernay, Cuisien (Ageian), MP10.
DISTRIBUTION. — Grauves, Cuis, Mancy, Monthelon, Mont Bernon, Chavot, St Agnan.
EMENDED DIAGNOSIS (modified from Savage et al. 1965). — Medium sized hippomorph (estimated skull length: 170-200 mm); well-developed lophodonty; hypoconulid linked to the middle of the hypolophid; cristid obliqua oriented lingually; lower molar lophs slightly notched.
DIFFERENTIAL DIAGNOSIS. — Much larger than all the other hippomorph species from the MP10 or earlier related localities of the Paris Basin. More lingual cristid obliqua than O. maldani , cingula generally more developed than O. maldani . More developed lophodonty than O. maldani .
DESCRIPTION
Teeth
The symphysis of the mandible is long (3.5 cm on MNHN-MA14902) and narrow (1.3 cm on MNHN-MA14902). The lower post-canine diastema is long (2.2 cm on MNHN-MA14902; 3 cm for MNHN-MA14908), but there is no post-p1 diastema ( Fig. 8I View FIG ). The p1 is simple and unicuspid. It can be uniradiculate (MNHN-MA14902, female morphotype, Fig. 8I View FIG ) or biradiculate (MNHN-CHO14801, MNHN-MA9-L, male morphotype, andFig. 8F, juvenile). The protoconid and metaconid of p2 are close one each other and obliquely aligned. A crest extends between the hypoconid and the labial side of the metaconid. The p3 is larger than the p2 but displays the same morphology. The protoconid and metaconid of p4 are close and almost aligned transversally. The entoconid is absent or well developed (MNHN-GR7572). A crest extends from the hypoconid to the labial side of the metaconid. The metaconid is twinned on MNHN-GR7572, and barely visible on most teeth due to the wear. The paralophid of m1-m2 is very short ( Fig. 8I, J, H View FIG ). The metaconid is twinned and the two cusps are aligned obliquely. The cristid obliqua joins the labial side of the metaconid. The hypoconulid is reduced and almost lingual. The postcristid is connected to the hypoconulid and to the hypolophid, at the level of the notch which lies closer to the entoconid. The cingulum is very reduced between protoconid and hypoconid, or absent. The paralophid of m3 is short, and the metaconid is twinned. The cristid obliqua joins the metaconid. The lobe of the hypoconulid is developed, it has the same length than the anterior part of the talonid, but stays narrower. The postcristid is linked to the center of the hypolophid, at mid-height. It forms an obliquely oriented horseshoe with the lingual part of the hypoconulid. The protolophid and the hypolophid are slightly notched.
The upper post-canine diastema is long (1.8 cm in MNHN-MA14948). There is no post-P1 diastema. The P1 is simple, elongated and posteriorly enlarged. One or two cusps are present labially. The P2 has three roots and the paracone and metacone are developed. Two morphologies of P2 are observable. In the first one, the P2 is triangular in occlusal view (MNHN-MA14948, Fig. 9J View FIG ), the protocone is shifted anteriorly and aligned transversely with the paracone, the paraconule and metaconule are absent, the protoloph is interrupted before joining the preparacrista, the parastyle is absent. The second morphology of P2 (MNHN-GR7400, Fig. 9A View FIG ; MNHN-MA14842, Fig. 9B View FIG ) displays a posteriorly enlarged premolar in occlusal view, with a posteriorly displaced protocone, which is aligned transversely with the metacone. The metacone is reduced sometimes missing ( Fig. 9A View FIG ). The protoloph is short and interrupted before joining the preparacrista. A cingular ridge extends posteriorly to the protocone. The parastyle is absent, but a small metastyle is present. The paraconule of P3 is small compared to the protocone and paracone, and the metaloph is located in the center of the tooth and interrupted labially at the base of the metacone. The parastyle of P4 is poorly developed ( Fig. 9D View FIG ). The metaconule is absent and the paraconule is wide. The metaloph (possibly the postprotocrista) is directed toward the middle of the two labial cusps, but preferably towards the metacone, and stops at the base of these cusps. The protoloph is interrupted before joining the preparacrista. The mesostyle of upper molars is generally absent ( Fig. 9I, J View FIG ), but it can be sometimes present as a small or developed crest running upward from the cingulum ( Fig. 9F, G View FIG ). The centrocrista is more or less labially flexed and high. The paraconule is larger than the metaconule, and both are well merged in the lophs. The cingulums are well developed but sometimes interrupted on the lingual side of the teeth. Generally, the M3 has a large enlargement of the posterior cingulum. A fusion of the posterior roots is observable on MNHN-CUI15513.
Deciduous teeth
The metaconule of DP3 is much larger than the paraconule ( Fig. 9E, H View FIG ). The lophs do not reach the labial cusps. The metaloph can form a crescent or is interrupted labially. The protoloph forms a crescent at its labial extremity. The parastyle is developed. The labial cingulum is interrupted close to the paracone. The DP4 is molariform, but the lingual side is narrower than the labial side ( Fig. 9H View FIG ). The centrocrista is flexed. The parastyle is slightly smaller than the paracone. It projects anteriorly and labially relative to the paracone. The metaconule is very small, and the paraconule is larger, but smaller than the protocone and paracone. The protoloph and the metaloph are well developed. The protoloph joins the preparacrista. The metaloph is interrupted at the base of the centrocrista, between the paracone and the metacone.
The dp2 metaconid is reduced and closely appressed to the protoconid ( Fig. 8F View FIG ). A crest extends anteriorly to the protoconid. A second crest runs from the protoconid to the posterior part of the tooth. The dp3 is molariforme ( Fig. 8C, F View FIG ). A small paraconid is present. The metaconid is twinned. The cristid obliqua joins the protolophid in the middle, slightly lingually. The hypoconulid is closely appressed to the base of the hypolophid, slightly lingually. The dp4 is molariform, and the morphology is similar to that of the molars ( Fig. 8D, F View FIG ). The tooth is slightly narrower at the trigonid, and longer than molars.
Postcranial elements
Humerus. Only the distal part is known. The supracondylar foramen (6 mm large and 5 mm high) perforates the radial fossa ( Fig. 9Q, R View FIG ). The ulnar fossa is triangular and perforated by the supracondylar foramen. The epicondyles are small, but the medial epicondyle projects farther than the lateral epicondyle. A small fovea is present below the medial epicondyle. A larger fovea is present on the lateral epicondyle.
Metacarpals. The metacarpal II is more robust than the metatarsals ( Fig. 9Y, Z View FIG ). The distal articular surface is asymmetrical and keeled. The distal articulation is asymmetric, with a medial part larger than the lateral part. The proximal facet is slightly concave. The trapezoid facet is oval and narrow.
Astragalus. The trochlea is deep and broad, slightly oblique with respect to the vertical axis of the astragalus ( Fig. 9K, L View FIG ). The trochlea does not join the navicular facet. The articular facets are well defined. The navicular facet is saddle-shaped. The ectal and sustentacular facets are well separated ( Fig. 9M, N View FIG ). The sustentacular facet joins the navicular facet. The distal calcaneal facet is laterally elongated.
Calcaneus. It is elongated and medio-laterally flattened ( Fig. 9O, P View FIG ). The calcaneal tuber is laterally compressed. The astragalar facet is small and slightly concave. The sustentacular facet is rounded, slightly triangular. The ectal facet is too worn to be described.
Metatarsals. The second metatarsal is slightly flattened mediolaterally on its proximal part ( Fig. 9S View FIG ). The distal facet was broken away. In dorsal view, the mesocuneiform facet has a concave crescent shape. The triangular entocuneiform facet is present posteriorly. The distal end curves medially away from metatarsal III. The metatarsal III is elongated and straight ( Fig. 9T View FIG ). The proximal cross section is triangular. The metatarsal III is flattened anteroposteriorly, and the distal cross section is elliptical. The proximal facet of articulation with the ectocuneiform is flat. The distal articular surface is symmetrical and keeled. The metatarsal IV is elongated and mediolaterally compressed proximally ( Fig. 9U View FIG ). This compression is less marked at the distal part. The distal articulation is asymmetric, with a medial part larger than the lateral part.
COMPARISONS
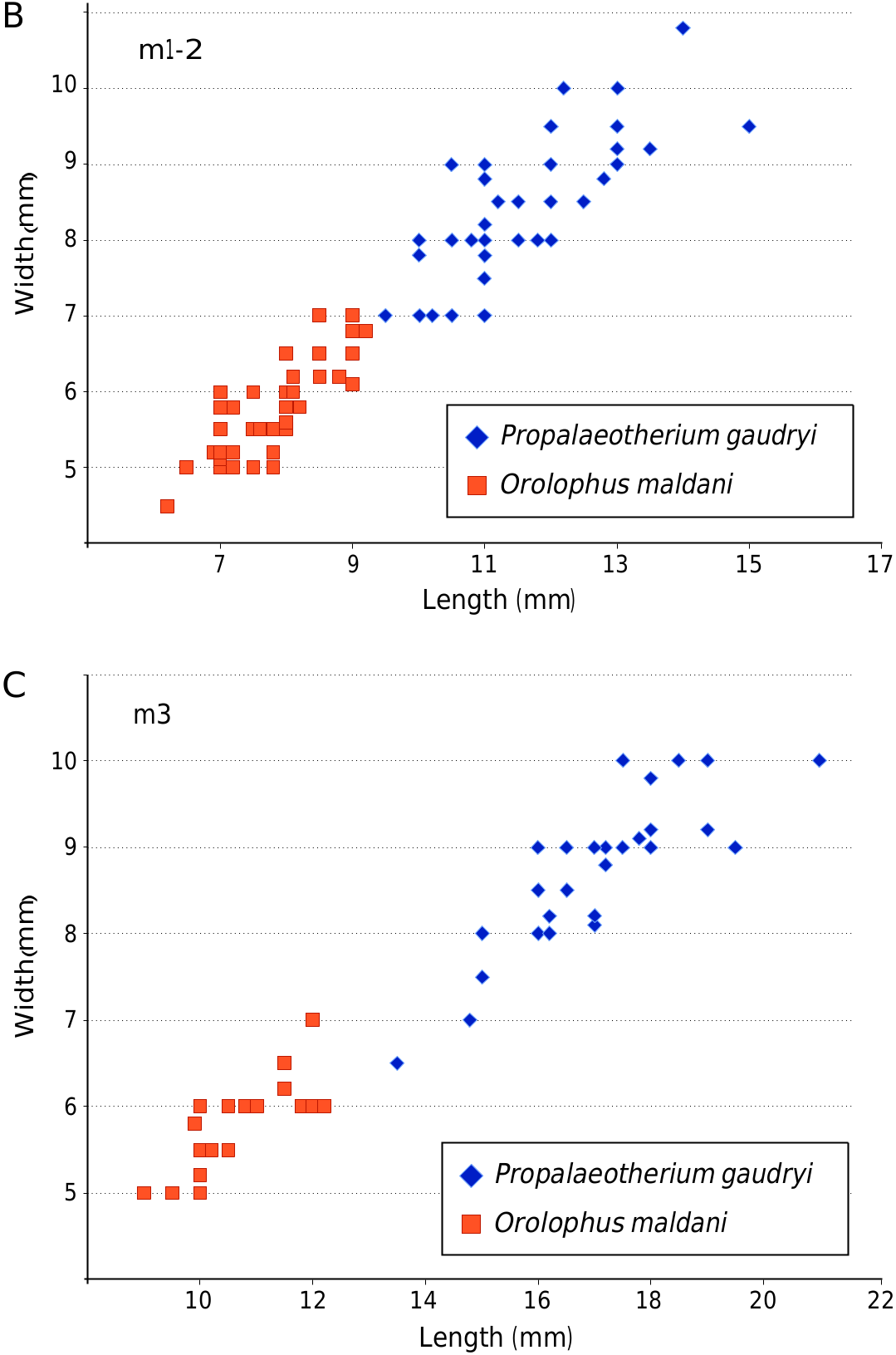
P. gaudryi is larger in size than Cymbalophus , Pliolophus , Hyracotherium , Orolophus and Hallensia . The teeth of P. gaudryi are more lophodont than those of O. maldani which is smaller in size ( Fig. 10 View FIG ). Their dental morphology is globally similar. The cristid obliqua of P. gaudryi is more lingual than that of O. maldani . Its cingulums are generally more developed than those of O. maldani . There is a wider range of size than other lower Eocene taxa. The upper molar mesostyle is variable, whereas it is always present in Propalaeotherium sudrei Remy, Krasovec & Marandat, 2016 and Propalaeotherium helveticum Savage, Russell & Louis, 1965 . The upper premolar mesostyle is absent in P. gaudryi but present in P. sudrei and Propalaeotherium helveticum . The parastyle is more projected anteriorly than in P. sudrei . P. gaudryi possesses a metaconule on upper molars, which is absent in Propalaeotherium isselanum Cuvier, 1824 .
COMMENT
Apart from the two morphotypes of P2, the dental morphology does not show significant differences. These two morphotypes of P2 are therefore attributed to intra-specific variability.
LECTOTYPE
Teilhard de Chardin (1921) designated the specimen MNHN-AL5210 as a lectotype. Unfortunately, this is a composite specimen whose two pieces were probably associated by Lemoine. The posterior part of the mandible does belong to Propalaeotherium gaudryi , but the anterior part (MNHN-AL6685) appears to be from a tapiroid (probably Chasmotherium [synonym of Hyrachyus ]) ( Savage et al. 1965). Specimen MNHN-AL5210 was therefore designated as a restricted lectotype by Savage et al. (1965).
SEXUAL DIMORPHISM
There is a great disparity in mandible height of the horizontal ramus and diastema length. There is also a great variation in mandible width at the level of incisors. MNHN-CHO14787 has three well-developed incisors on each side, while those of MNHN-MA14902 are narrower and its alveolus indicates a very small I3. The same morphotypes as in P. eulaliensis Danilo, Remy, Vianey-Liaud, Marandat, Sudre & Lihoreau, 2013 ( Danilo et al. 2013), Pliolophus quesnoyensis ( Bronnert et al. 2018) and Lophiodontidae ( Vautrin et al. 2019) are observable: variation in the height of the mandible and the angle of the vertical branch. Mandibles with a high horizontal ramus and a vertical ascending branch are assigned to males ( Fig. 9L View FIG ), mandibles with a low horizontal ramus and an inclined vertical branch are assigned to females ( Fig. 9K, M View FIG ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Propalaeotherium gaudryi ( Lemoine, 1878 )
| Bronnert, Constance & Métais, Grégoire 2023 |
Propalaeotherium gaudryi
| REMY J. A. 2017: 290 |
Propachynolophus gaudryi
| SAVAGE D. E. 1965: 15 |
| TEILHARD DE CHARDIN P. 1921: 71 |
Pachynolophus (Propachynolophus) gaudryi
| LEMOINE V. 1891: 285 |