
Hallensia louisi Hooker, 1994
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2023v45a9 |
|
publication LSID |
urn:lsid:zoobank.org:pub:1C430978-5EE6-49AE-AF7C-23C710161CB7 |
|
DOI |
https://doi.org/10.5281/zenodo.8043653 |
|
persistent identifier |
https://treatment.plazi.org/id/D066B24B-521B-B67D-FC32-FE6EDF6FFD85 |
|
treatment provided by |
Felipe |
|
scientific name |
Hallensia louisi Hooker, 1994 |
| status |
|
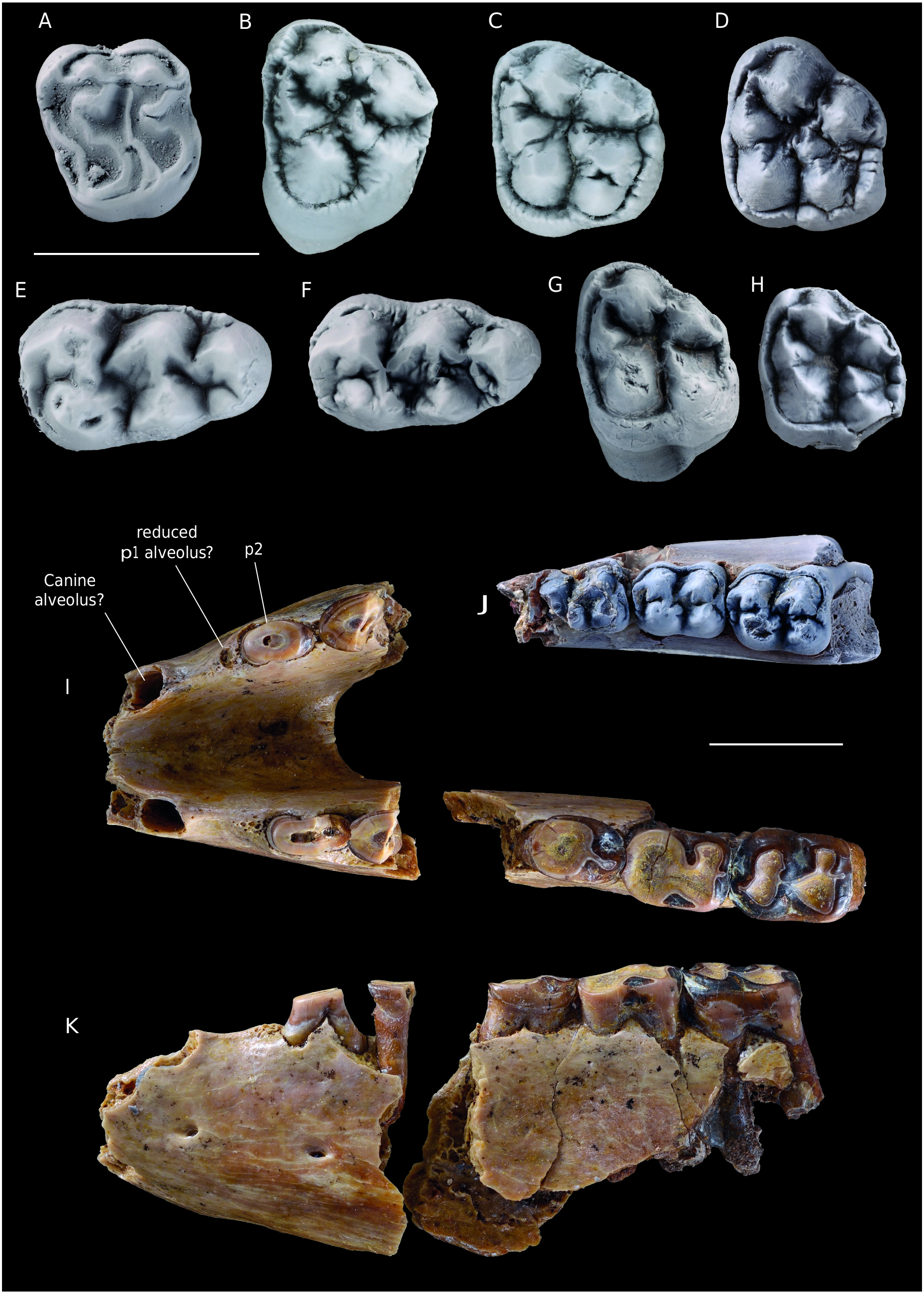
( Fig. 15 View FIG )
Hallensia louisi Hooker, 1994: 49 .
Propachynolophus sp. – Teilhard de Chardin 1921: 69, fig. 33C, pl. 3, fig. 30.
Hyracotherium [sic] – Teilhard de Chardin 1921: 52, fig. 26B. — Savage et al. 1965: 6-8, 10, 11, 13, figs 2d, f, i, j, 4f-h.
HOLOTYPE. — MNHN-MU218-L , left M1/2 .
PARATYPES. — MNHN-MU12371, right M3; MNHN-MU201-L, left M3; MNHN-MU221-L, left M3; MNHN-MU12303, right mandible with dp4-m2; MNHN-MU6283, right m1/2; MNHN-MU12391, right m1/2, broken .
MATERIAL. — Avenay: Fragment of dentary and symphysis (L: MNHN-AV-841-Ph), m3 (L: MNHN-AV65519), M1/2 (R: MNHN-AV4770).
Condé-en-Brie: DP4 (R: MNHN-CB1588; L: MNHN-CB1574); M1/2 (L: MNHN-CB216, MCB0132, CB217 [broken]); M3 (R: MNHN-CB1568; L: MNHN-CB1560); m2 (L: MNHN-CB668); m3 (R: MNHN-CB4-CA; L: MNHN-CB224).
Gland: DP4 (R: MNHN-GLD216-L); M1/2 (R: MNHN-GLD281- L).
Grauves: M3 (R: MNHN-GR7574).
Oosterzele: p4 (R: IRSNB.M 1862); m3 (L: IRSNB.M 1861) ( Smith et al. 2004) .
TYPE LOCALITY. — Mutigny (Marne, France), MP8+9.
DISTRIBUTION. —Mutigny (Marne, France), Condé-en-Brie (Aisne, France), Avenay (Marne, France), Gland, Oosterzele ( Belgium) (MP8+9); Grauves (Marne, France) (MP10).
EMENDED DIAGNOSIS. — Small species of Hallensia ; small upper molar parastyle; m3 hypoconulid linked to the hypoconid; upper molar centrocrista more or less flexed, sometimes with a mesostyle; large cusps, with a “bloated” appearance.
DIFFERENTIAL DIAGNOSIS. — Smaller than H. parisiensis and H. matthesi ; more bunodont than Cymbalophus , Pliolophus , Hyracotherium and Orolophus ; differs from Orolophus by a hypoconulid linked to the hypoconid. Mandibular symphysis shorter and larger than in Cymbalophus and Pliolophus .
DESCRIPTION
Teeth
The upper molars are bunodont or sub-lophodont, with ( Fig. 15B View FIG ) or without mesostyle ( Fig. 15C, D View FIG ). The parastyle is reduced, the cingulum is wider posteriorly. The metaloph is continuous or interrupted at the base of the metacone. The conules are developed and close to lingual cusps. The protocone is the largest cusp. The cingulum is crenellated, sometimes forming small cuspids.
The mandibular symphysis is short and broad ( Fig. 15I, K View FIG ). It extends posterior to the limit p2-p3. A diastema is present anterior to the p2, up to the large alveoli which probably correspond to canine alveoli. The most anterior cheek tooth still present is presumed to be a p2. Its occlusal outline is elongated oval, and the two roots are widely separated. This corresponds to the morphology generally observed for a p2, the p1 usually has fused roots. The shape of the bone seems to indicate that resorbed alveoli, which could correspond to the alveoli of the p1, are present just ahead of the p2. The large alveoli situated anteriorly are deep and vertical, which is probably the alveoli of canines.
The m1-2 have a quite bunodont appearance, the cusps are very rounded ( Fig. 15J View FIG ). The paralophid is very short. The metaconid is twinned. The protolophid is notched. The hypolophid is either absent or very reduced. The hypoconulid is linked to the hypoconid by a postcristid when the hypolophid is absent; it is attached to the hypolophid when the latter is present. The cristid obliqua is poorly developed, it is oriented toward the base of the protolophid, in the middle. The labial cingulum is continuous. The labial sides of the protoconid and the hypoconid are very inclined (around 45°), whereas the lingual side of these cuspids is almost vertical. The m3 is wide and bunodont ( Fig. 15E, F View FIG ). The paralophid is quite elongated. The metaconid is twinned. The protolophid and the hypolophid are moderately developed. The cristid obliqua is oriented toward the protolophid almost in the center of the tooth, slightly labially. The labial cingulum is continuous. The hypoconid is linked to the hypoconulid by a postcristid which is lingually flexed. A small cusp is sometimes present on the lingual side of the hypoconulid.
Deciduous teeth
The DP4 is molarized and narrower posteriorly ( Fig. 15A View FIG ). The centrocrista is straight. The paraconule and the metaconule are developed. The parastyle is small. The metaloph ends with a metacone fold.
The dp4 seems to be molarized, and a wide entoconid is present ( Fig. 15J View FIG ). The labial wall of the cuspids is very steep. A small hypolophid seems to be present. The lingual and labial cusps are obliquely aligned. The cristid obliqua is oriented low, toward the center of the protolophid.
COMPARISONS
The teeth are more bunodont than in other early Eocene perissodactyls. The species is smaller than H. parisiensis and H. matthesi . The junction of the lower molar hypoconulid is similar to that of Pliolophus quesnoyensis and Pliolophus barnesi . A small mesostyle is sometimes present, as in Orolophus maldani and Hyracotherium . The mandibular symphysis is shorter and more robust than in Pliolophus , Cymbalophus and P. gaudryi .
COMMENT
Hallensia was first identified as a phenacodontid condylarth ( Franzen & Haubold 1986). The discovery of a complete skeleton in Messel ( Germany) revealed that Hallensia displays a saddle-shaped navicular facet of the astragalus which led Franzen (1990) to place it within perissodactyls. Bajpai et al. (2006) considered Hallensia as close to the Indian cambaytheres, which they considered as a perissodactyl family. Indeed, the genus Hallensia has many similarities with the cambaytheres, recently described in India ( Bajpai et al. 2006; Cooper et al. 2014; Rose et al. 2014). They notably have very bunodont teeth and a quasi-absence of lophs. The parastyles are very small or absent and the cingula are generally well developed. They mainly display the synapomorphy of the perissodactyls on the astragalus. Cambaytherium , however, has features found in phenacodonts but absent in perissodactyls and Hallensia , such as a developed deltopectoral crest and five metapodials per limb ( Rose et al. 2014).
Hallensia also shows tooth wear that forms horizontal facets, as in Cambaytherium . This wear pattern is sometimes very pronounced ( Fig. 15I View FIG ). The bunodont morphology of the teeth, as well as this important wear pattern indicates a diet based on hard food. This is confirmed by the discovery of fine sand in the stomach contents of H. matthesi , as well as remnants of stems and few leaf remains ( Franzen 1990). It probably had to feed on stems close to the ground, which explains the ingestion of clastic material such as sand.
The mandible attributed to H. louisi has very worn teeth. Hooker (1994) indicates that the anterior alveoli are those of p1. The shape of the bone, slightly “fenestrated” in front of the p2 seems however to indicate the presence of resorbed alveoli, which would correspond to the p1. In addition, the broad and deep shape of the alveoli is reminiscent to that of the canines of Hallensia matthesi , which are positioned quite vertically compared to European hippomorphs ( Cymbalophus cuniculus , Pliolophus quesnoyensis ).
The size of Hallensia increases through time. Hallensia louisi (MP8-9, 10) is the oldest species and is very small, and H. parisiensis (MP8-9, 10), which first appeared slightly later has an intermediate size between H. matthesi and H. louisi . The largest Hallensia species, H. matthesi (MP11-13), is similar in size to Cambaytherium thewissi and smaller than Cambaytherium bidens .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hallensia louisi Hooker, 1994
| Bronnert, Constance & Métais, Grégoire 2023 |
Hallensia louisi Hooker, 1994: 49
| HOOKER J. J. 1994: 49 |
Propachynolophus sp.
| TEILHARD DE CHARDIN P. 1921: 69 |
Hyracotherium
| SAVAGE D. E. & RUSSELL D. E. & LOUIS P. 1965: 6 |
| TEILHARD DE CHARDIN P. 1921: 52 |