
Onychiurus gevi, Arbea, Javier, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.209507 |
|
publication LSID |
lsid:zoobank.org:pub:ED4A1E9E-115F-489B-92A7-89710819715E |
|
DOI |
https://doi.org/10.5281/zenodo.6180617 |
|
persistent identifier |
https://treatment.plazi.org/id/D07B87D8-FFF2-0479-CBAF-7F9267025C78 |
|
treatment provided by |
Plazi |
|
scientific name |
Onychiurus gevi |
| status |
sp. nov. |
Onychiurus gevi sp. nov.
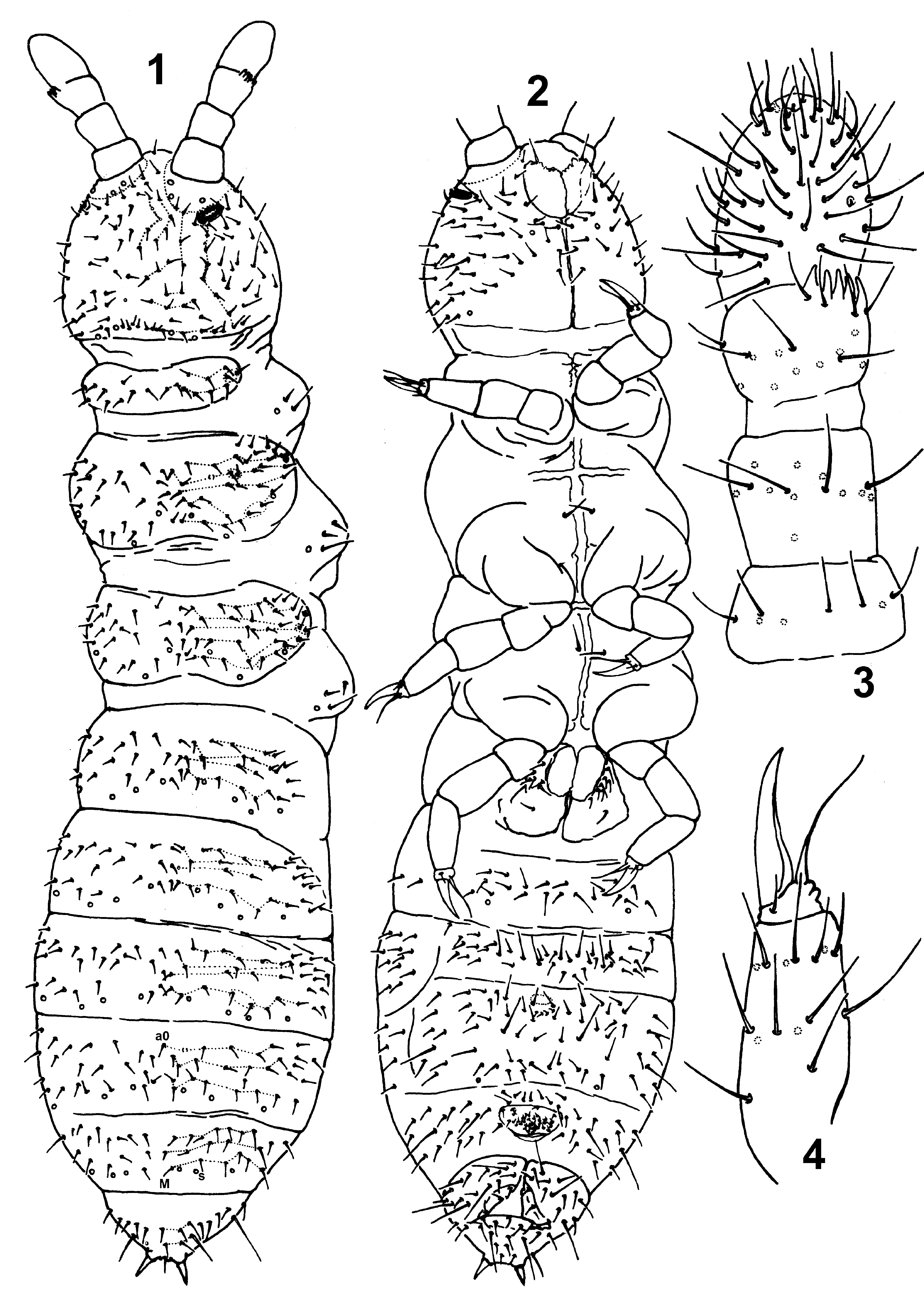
Figs. 1–12 View FIGURES 1 – 4 View FIGURES 5 – 12
Type locality. Spain, Sima Gesm, Coordinates 36º41’32’’N, 5º00’22’’W, 1752 m a.s.l. in the Sierra de las Nieves karst of Tólox (Province of Málaga).
Type material. Holotype: female (mounted on slide labelled EML0017): Spain, Málaga, Tólox, Sierra de las Nieves karst, Sima Gesm, 27.VIII.2006, G.E.V. (Speleological Club of Villacarrillo) leg. Paratypes: one reproductive male, seven females (mounted on slides labelled EML0017). Holotype and paratypes deposited in The Museum of Zoology at Pamplona, University of Navarra.
Other material. One female (mounted on slide labelled EML0016): Spain, Málaga, Tólox, Sierra de las Nieves karst, Sima Gesm, 15.VII.2006, G.E.V. leg. Two males and three females (mounted on slides labelled EML0034): Spain, Málaga, Tólox, Sierra de las Nieves karst, Sima de Raja Helada (TO-8), Coordinates 36º41’35’’N, 5º00’27’’W, 1747 m a.s.l., 12.X.2009, Patricia Carrasco leg. All specimens deposited in the private collection of the author.
Etymology. The species is dedicated to the members of the Speleological group of Villacarrillo (G.E.V.), who collected specimens of this species from Málaga caves.
Description. Colour white in alcohol. Body length (excluding antennae) of adults: holotype 2.8 mm, reproductive males 1.77–2.29 mm, reproductive females 2.27–3.02 mm. Cuticle granulation more or less uniform and fine, somewhat coarser on terga, head capsule, and around anal spines.
Antennal bases well marked. Antennae slightly shorter than head; ratio antennae/head diagonal = 0.86. Ant. IV with subapical organite; microsensillum in latero-external position, approximately two-fifths length from the base; sensilla not well distinguishable from ordinary chaetae; invaginated apical bulb small. Ant. I, II and III with 8, 14 and 17 chaetae respectively. AIIIO as in Fig. 5 View FIGURES 5 – 12 , with five papillae, two small sensory rods, two bent and smooth sensory clubs, four guard chaetae, and lateral microsensillum.
PAO consists of 16–17 finely granulated vesicles ( Fig. 6 View FIGURES 5 – 12 ). Labrum as in Fig. 7 View FIGURES 5 – 12 with 5,4,2 chaetae. Labial palp of type A with five proximal chaetae; labial papillae A, B, C, D and E with 1, 4, 0, 3 and 3 guard chaetae respectively ( Fig. 8 View FIGURES 5 – 12 ); chaetotaxy of the basomedian field (submentum) with 4+4 chaetae ( Fig. 8 View FIGURES 5 – 12 ), basolateral field (mentum) with 5 chaetae ( Fig. 8 View FIGURES 5 – 12 ); outer maxillary lobe with one basal chaeta and without sublobal hairs ( Fig. 8 View FIGURES 5 – 12 ). Mandible with strong molar plate and four (rarely five) apical teeth as in Fig. 9 View FIGURES 5 – 12 ; maxilla bearing three teeth and six lamellae as in Fig. 10 View FIGURES 5 – 12 .
Dorsal pseudocellar formula 32/033/44433, ventral 11/000/0101; all subcoxa I with one pseudocellus. Parapseudocelli not visible on the body.
Dorsal chaetotaxy as in Fig. 1 View FIGURES 1 – 4 and Tables 2–4 View TABLE 2 View TABLE 3 View TABLE 4 ; nearly symmetrical, poorly differentiated into meso-, and microchaetae, sometimes extra chaetae and asymmetries have been observed; macrochaetae only differentiated on the two last Abd. tergites. Th. II and III with microsensilla located laterally. Body sensilla cylindrical, poorly differentiated. Head with unpaired dorsal chaeta d0. Th. I with 10–11+10–11 chaetae. Th. II to Abd. III with 4,4,3,3,3 chaetae respectively on both sides of axial line and without unpaired axial chaetae. Abd. IV and V without unpaired axial chaetae; rarely chaeta a0 present on Abd. IV. Ratio chaetae M/s = 1.4–1.5 on Abd. V. Abd. VI with 2–3 unpaired axial chaetae: (a0), m0 and p0 ( Fig.11 View FIGURES 5 – 12 ). Subcoxa I of I, II, III pairs of legs with 2,3,3 chaetae respectively. Anal spines rather short, about 0,6 of inner edge of the claw III length.
g 9 chaetae; g4 absent
Chaeta number/serie 1 2 3 4 5 6 7 Th I
m - - m - m - m Ventral body chaetotaxy as in Fig. 2 View FIGURES 1 – 4 , with 0+0, 1+1 and 1+1 ventral chaetae on pro-, meso- and metathorax respectively. VT with 7–8+7–8 apical chaetae and 1+1 basal chaetae. Furca reduced to a finely granulated area, with 2+2 posterior setulae arranged in one row ( Fig. 12 View FIGURES 5 – 12 ). Male ventral organ absent.
Tibiotarsi I, II, and III with 15–16(7,7–8,1), 15–16(7,7–8,1) and 15(7,7,1) chaetae respectively; distal whorl with seven chaetae ( Fig. 4 View FIGURES 1 – 4 ). Claws without teeth. Empodial appendage slender, as long as inner edge of a claw, without basal lamella ( Fig. 4 View FIGURES 1 – 4 ) (appendage length 1.0–1.1 inner edge of claw).
Ecology. O. gevi sp. nov. is bisexual species. It has been collected in cold and wet cave habitats. It has been found together with Deuteraphorura cebennaria ( Gisin, 1956) ( Arbea et al. 2011) .
Discussion. O. gevi sp. nov. resembles the Onychiurus obsiones species group sensu Kaprus’ (2008). It clearly differs from all members of the group in size (1.7–3.0 in O. gevi sp. nov. versus 0.6–1.0 mm), number of pseudocelli on Abd. tergites I–III (4,4,4 in O. gevi sp. nov. versus 3,3,3), ventral pseudocellar formula (2/000/0101 in O. gevi sp. nov. versus 2/000/0001), and habitat preferences (caves in Málaga, S Spain in O. gevi sp. nov. versus soil in mountains beech forest in Poland, and sandy habitats in arid and semiarid regions of N Africa, SE Europe, Minor and Middle Asia) (see Kaprus’ 2008 and Smolis & Skarżyński 2009).
Some Onychiurus species: O. boldorii , O. nathanieli , O. paoletti , O. stillicidii also share the same dorsal pseudocellar formula on head and Th. as the new species, but differ in dorsal pseudocellar formula on Abd. (4,4,4,3,3 in O. gevi sp. nov. versus 3,3,3,4, 3 in O. boldorii and O. stillicidii , 3,3,3,5,3– 4 in O. nathanieli , and 3,3,3,5, 3 in O. paoletti ) and ventral pseudocellar formula (see Table 5).
The new species is distinguished from all other species of the genus by the number of pseudocelli on Abd. I–III (see Table 5).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.