
Erenna sirena, Pugh, P. R. & Haddock, S. H. D., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4189.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:0F0EE4C3-05B9-4248-944D-B41DC8361CB2 |
|
DOI |
https://doi.org/10.5281/zenodo.6067134 |
|
persistent identifier |
https://treatment.plazi.org/id/D277A579-4C3E-296B-FF68-897BF7E3E247 |
|
treatment provided by |
Plazi |
|
scientific name |
Erenna sirena |
| status |
sp. nov. |
Erenna sirena View in CoL sp. nov.
Diagnosis. Flattened nectophores with pairs of upper lateral, lower lateral and vertical lateral ridges. The upper laterals do not divide and end at some distance above the ostium. Two types of bract; the Type A being distinguished by their relatively small size and absence of any transverse ridges or flaps. Very characteristic tentilla where the terminal process is modified to form a red bioluminescent appendage that acts as a lure.
Material examined. Thirteen specimens of Erenna sirena sp. nov. are known to have been collected and six of these have been examined for the present description, namely:
ROV Dive Date Position Depth (m) Tiburon 594-SS7 17 July 2003 36°12.99'N 122°32.00'W 1935 Tiburon 678-D5 25 May 2004 35°28.84'N 123°52.74'W 2428 Tiburon 681-D3 27 May 2004 36°33.81'N 122°11.65'W 2262 Tiburon 981-D3 11 May 2006 36°36.13'N 122°22.57'W 2188 Tiburon 1040-D3 1 October 2006 34°17.23'N 124°03.10'W 2061 Tiburon 1154-SS1 28 November 2007 35°49.97'N 122°39.99'W 1920 Holotype: The specimen from Tiburon Dive 1154-SS1 has been designated the holotype and will be donated to National Museum of Natural History, Smithsonian Institution, Washington, D.C., U.S.A.
Description. In general the in situ specimens were up to c. 70 cm in length, with the siphosome usually slightly longer than the nectosome ( Figure 37 View FIGURE 37 ). The cell patches, including nematocysts, at the distal ends of the bracts were prominent, but would not serve to identify the species in situ. However, the thick clusters of tentilla were quite distinctive, and absolutely so if they could be seen in close-up so that the red lures could be observed ( Figure 38 View FIGURE 38 ).
Pneumatophore. The preserved, and partially exploded, pneumatophore measured c. 3.5 mm in length and 1.75 mm in diameter, and was completely featureless.
Nectosome: Up to eighteen pairs of mature nectophores were observed on in situ specimens.
Nectophores: There were 26 detached mature and three young nectophores included with the type specimen, and one nectophoral bud. These nectophores measured up to 30 mm in width and length. One of the distinctive characters of the very young nectophores ( Figure 39 View FIGURE 39 A) was the fact that the upper lateral ridges ended, distally, well above ostial level and without dividing. Both the upper and lower lateral ridges were well-defined while the vertical laterals were less so, although they clearly joined with the other two ridges. There was a prominent lateral spot on either side of the nectophore just distal to the vertical lateral ridges and as the lateral radial canals passed over the lateral apices of the nectosac they were considerably thickened. The lateral ostial processes were not prominent at this stage, and the canal running through them was ill-defined. However, they bore nematocysts measuring c. 90 x 25 µm. There also was a prominent patch of ectodermal cells on the upper side of the ostium, where the upper canal joined the ring canal.
As the nectophores enlarged, the muscle-free zone on the proximal part of the lower side of the nectosac became apparent, and the thrust block began to develop and, at that stage, consisted of two small lobes ( Figure 39 View FIGURE 39 B, C).
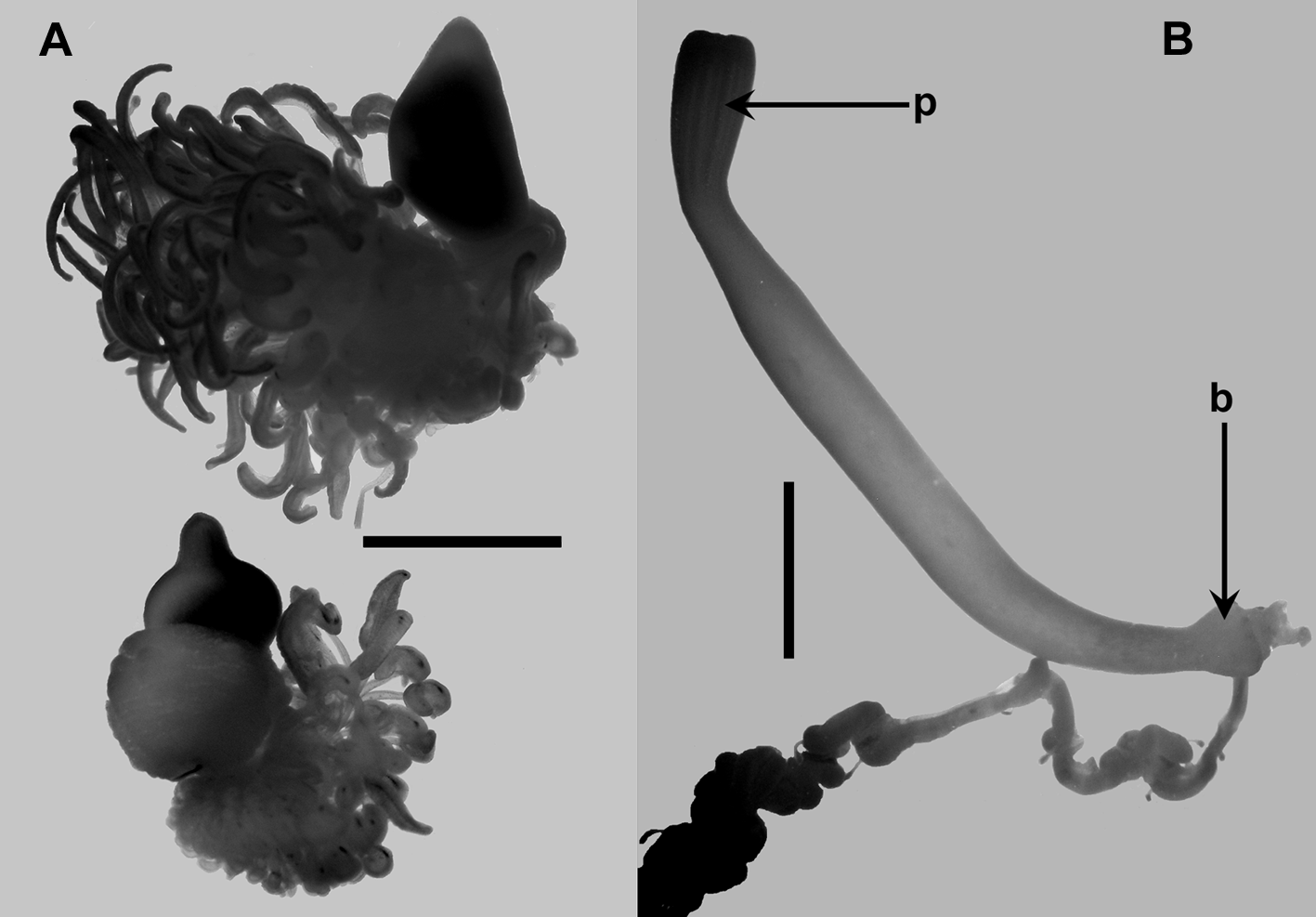
The fully developed nectophores ( Figure 40 View FIGURE 40 ) had the typical Erenna form, i.e. compressed in the upper/lower plane and with a distinct triangular lateral facet in the proximal half of the nectophore, delimited by the upper, lower and vertical lateral ridges. Distally, the upper lateral ridges turned inwards and ran directly toward the midline, before turning back toward the ostium and becoming much less distinct and gradually petering out. The lower ones continued to the ostium. Just distal to the vertical lateral ridge there was a small patch of ectodermal cells. The axial wings increased further in size and formed a pair of extensive lobes. The thrust block remained relatively small but now took on its characteristic form, consisting of two rounded lobes, on the upper side of the nectophore, behind which where two smaller digitate flaps.
The nectosac occupied the distal half of the nectophore and had a distinct muscle-free zone at it proximal end. The ascending mantle canal was quite short and gave rise to a long pedicular canal that ran to the lower wall of the nectosac just below its apex. It gave rise directly to all four radial canals, the upper and lower running directly to the ostium, while the laterals followed the contours of the lateral walls of the nectosac. Two long, but narrow lateral ostial processes extended out from the ostium.
Siphosome: A very unusual, if not unique, situation was noted on the siphosome of Tiburon Dive 1040 specimen observed in situ ( Figure 41 View FIGURE 41 ). Here the tentacles, particularly toward the posterior end of the colony, appeared to possess long contractile pedicles, such that the tentacle itself was hanging loose at some distance from the siphosomal stem.
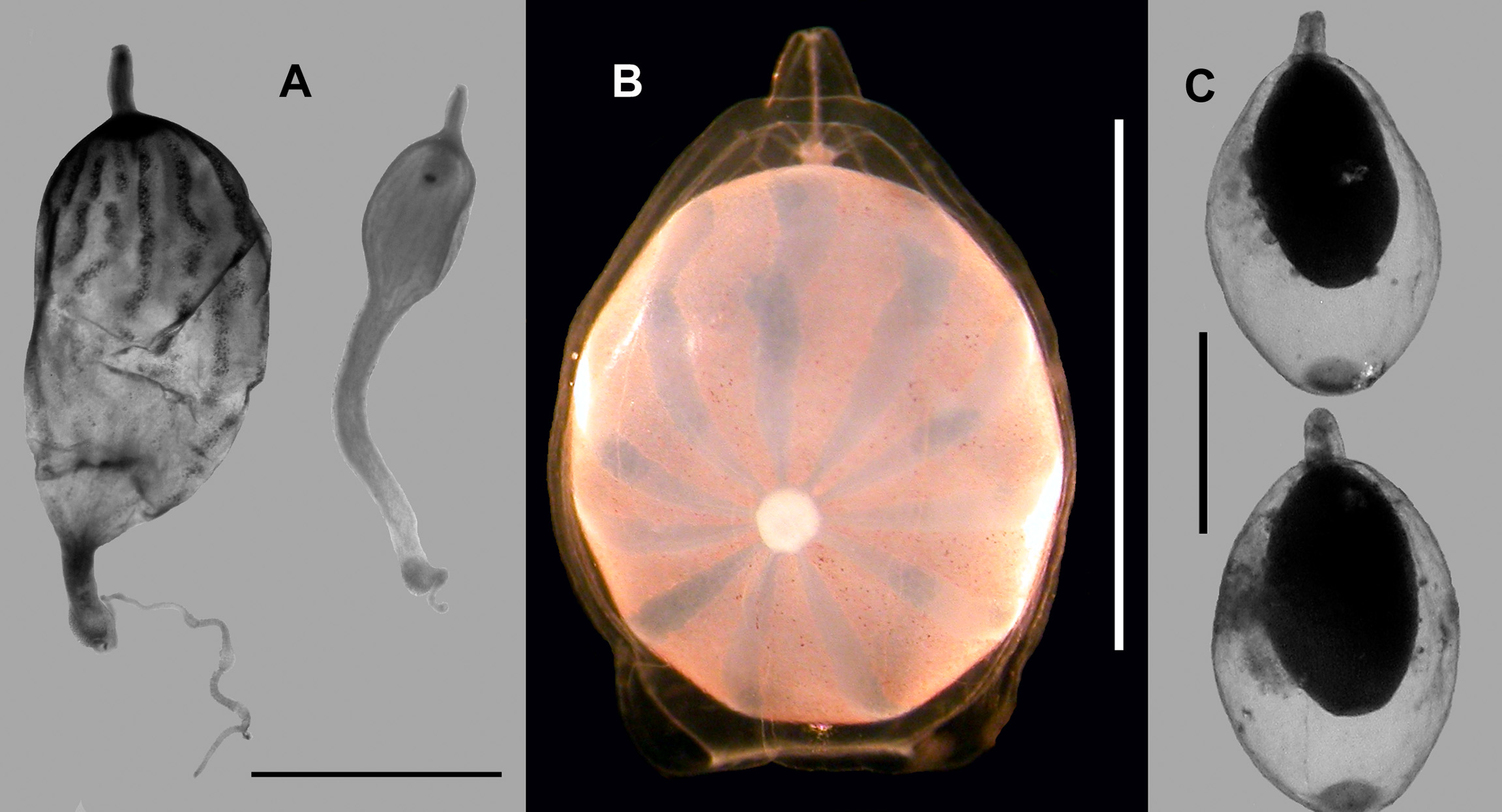
Bracts: Typically there were two types of bracts, with the Type A ( Figure 42A View FIGURE 42. A ) being longer and thinner, and the Type B ( Figure 42 View FIGURE 42. A B) shorter and fatter. The Type A bracts measure up to 27 mm in length and 14 mm in maximum width. They possessed two pairs of lateral cusps, with the distal pair being almost symmetrical, while the proximal pair were distinctly asymmetrical, with the cusp on the inner side being considerably more distal than the other. The distal end of the mature bracts was pointed and formed a right-angle. Proximally the inner side was thickened. The bracteal canal arose on the upper side of the bract and, once it had passed over to the lower side, remained in contact with the lower wall of the bract for most of its distance. However, distally it narrowed considerably and ran obliquely through the mesogloea to end below a large patch of cells, including nematocysts, that was raised above the upper surface of the bract just prior to its distal end. In the youngest bracts, this cluster of cells was formed into a ball that projected beyond the distal tip of the bract. A small circular patch of ectodermal cells was present in the mid-line on the upper side of the bract, usually approximately on a level with the more distal of the proximal pair of lateral cusps. This was not always to be seen on the younger bracts.
The nematocysts ( Figure 42 View FIGURE 42. A C) in these clusters were of one type, although somewhat variable in size ranging from 90–103 µm in length and 25–30 µm in diameter, and presumably were the same as those on the lateral ostial processes of the nectophore.
The Type B bracts ( Figure 42 View FIGURE 42. A B) measured up 20 mm in length and 16 mm in width. They also possessed two pairs of lateral cusps, but the proximal pair was much closer to being symmetrical than those on the Type A bracts. The course of the bracteal canal was the same as for the Type A bract, as was the proximal thickening on the inner side of the bract itself. The distal patch of nematocysts, etc, on the upper side of the bract, was similarly raised above the surface, but generally smaller in size, especially in the mature bracts where the number of nematocysts was reduced, either by erosion or usage.
Gastrozooid and tentacle: The younger gastrozooids ( Figure 43 View FIGURE 43 A) had a relatively small stomach and proboscis region and a large basigaster, to which the young tentacle with developing tentilla was attached. The mature gastrozooids ( Figure 43 View FIGURE 43 B) often were more elongate, up to 3 cm in length, with a small slightly inflated basigaster, a long stomach region, and a narrow proboscis within which the so-called hepatic stripes were apparent.
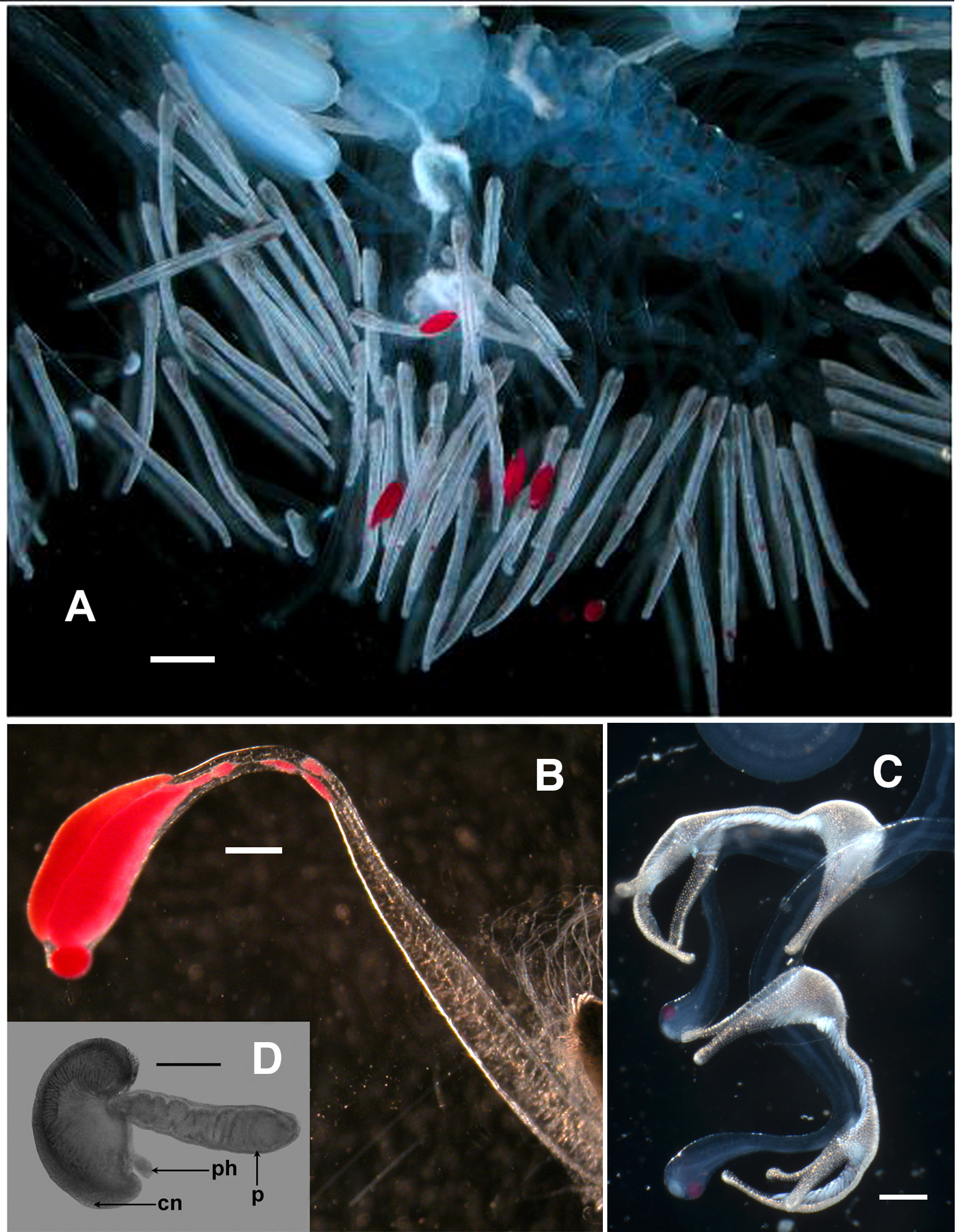
Each tentacle bore a considerable number of tentilla ( Figures 37 View FIGURE 37 , 44 View FIGURE 44 A) at various stages of development ( Figures 44 View FIGURE 44 B, D, 45). Although, for many, the cnidoband was of about the same length, the degree to which the lure was developed varied considerably. Only a few had the fully developed red-pigmented lure ( Figure 44 View FIGURE 44 A), while in many the lure remained small and whitish in colouration. As Haddock et al. (2005) showed the milky-white appearance of the younger lures was due to the presence of bioluminescent material that later became covered in a red fluorescent material with multimodal emissions. The relative sizes of the cnidoband and the lure varied during the development of the tentillum. At the very youngest stage ( Figure 44 View FIGURE 44 D), in its preserved state, it consisted of a highly contracted pedicle, with the relatively wide gastrovascular canal concertinaed. The cnidoband formed a hemispherical partial covering to an inflated cavity, with the very small stalkless photophore projecting out from the other side. The earliest stage photographed in life ( Figure 45 View FIGURE 45 A), however, showed the milky-white bioluminescent lure, again without a stalk, to be considerably larger than the ill-developed cnidoband. As development progressed ( Figure 45 View FIGURE 45 B–E) the size of the cnidoband increased rapidly, while that of the photophore remained the same or even slightly decreased. At first ( Figure 45 View FIGURE 45 B–C) it developed a short free stalk, but later this seems to have been resorbed ( Figure 45 View FIGURE 45 E). Only at the final stages in the development of the tentillum did the photophore, with its covering of fluorescent material, greatly increase in size and the vacuolated stalk considerably elongate. At all stages of development the tip of the stalk could be flicked through strong contractions of the stalk itself. One young specimen, with only sixteen nectophores, was found to have retained the larval tentacle at the posterior end of the siphosome ( Figure 44 View FIGURE 44 C). The eight attached tentilla had a unique "anteater-shaped" cnidoband and the lure was more bulbous-shaped than a typical mature lure. It was flicked in a similar manner to the adult one, through contractions of the stalk, and the photophore consisted of a small hemispherical protuberance from close to its distal end.
The bioluminescent and fluorescent properties of the lure have been dealt with by Haddock et al. (2005) and further details can be found on the web. One of these, http://www3.mbari.org/news/news_releases/2005/lures.html, contains a figure showing the red distal tips of the lures, with an inset showing the side view of one of these, which was considered closely to resemble a copepod..
The usual three types of nematocyst were found on the cnidoband ( Figure 46 View FIGURE 46 ). On each side there were several rows of, what were presumed to be, large microbasic anisorhizas ( Figure 46 View FIGURE 46 C) that measured c. 135 x 21 µm before discharge, but were longer after discharge as the capsule straightened out. Very occasionally, a smaller version of this type of nematocyst, which measured c. 85 x 25 µm, was observed. The bulk of the cnidoband consisted of two, smaller types of nematocyst. A cylindrical form ( Figure 46 View FIGURE 46 A), whose tubule was without a shaft and possibly atrichous, had a capsule that measured c. 39 x 12 µm; and a pear- shaped form ( Figure 46 View FIGURE 46 B) that measured c. 31 µm in length and 15 µm in maximum diameter. The latter were considerably more abundant than the former and, when discharged, they showed the peculiar asymmetrical proximal swelling with a long tubule extending from it that has been described above for Erenna richardi .
Palpon: The palpons ( Figure 47 View FIGURE 47 A) were very similar in morphology to those described above for Erenna insidiator . The mature ones had a short pedicle, to which the palpacle was attached, that gradually expanded into the main body region, covered in an array of long irregular patches of large ectodermal cells interspersed with or overlaying more homogeneous smaller cells, and then rapidly narrowing into the long distal proboscis. The largest palpons measured c. 15 x 5 mm. Unlike those of E. insidiator no clusters of elongate nematocysts were found around the distal mouth region. Occasional ones were observed, but it was not clear if these were definitely positioned there or were being ingested or egested.
Gonophore: The female gonophores were of a considerable size ( Figures 38 View FIGURE 38 & 47 View FIGURE 47 B) and, in their preserved state, measured c. 6 mm in length and 4.5 mm in diameter. They had a short triangular pedicle through which passed the pedicular canal. On reaching the subumbrella this canal initially gave rise to five radial canals and, after a short distance, three of these divided so that eight canals ran down to the ostial ring canal. Whether this was also the case for the male gonophores could not be elucidated. After preservation the egg turned a dark brown colour and no superficial structures could be seen. However, in life there were 11 bluish ribbon-like structures which connected with the region that was presumed to contain the nucleus ( Figure 49 View FIGURE 49 B). Female gonophores at all stages of development were found on the colonies.
The male gonophores ( Figure 47 View FIGURE 47 C, 48–49) were borne on elongate gonodendra that were only developed on the older, more posterior, cormidia. Each gonophore ( Figure 47 View FIGURE 47 C) measured c. 2.1 mm in length and 1.25 mm in diameter, including a short narrow pedicle. As the gonophores matured the sexual products increased in volume so as to completely filled the subumbrella cavity, and the mouth opening broke through and widened considerably. No nematocysts were found around the opening.
Remarks. The tentilla of Erenna sirena , although resembling those of E. laciniata , as noted above, more closely resemble those of Parerenna emilyae (see Pugh, 2001), particularly in the presence of what appears to be a lure, although the colour of that of P. emilyae is unknown. Since all the other species Erenna have highly distinctive tentilla it is reasonable to examine the possibility of these species might be one and the same. The original material of P. emilyae was not in pristine condition, having been examined for bioluminescence before preservation, and indeed has deteriorated further since it was originally described, through poor curation. However, there are striking differences in the relative size of the various zooids of the two species. For, instance, the nine nectophores of P. emilyae , which appeared to be mature, had maximal dimensions of 13 mm in length and breadth. That compares with the 30 x 30 mm, respectively, for E. sirena . Indeed, a youngish nectophore of the latter, with typically thickened lateral radial canals, was larger than any of those of P. emil yae . The young tentilla of E. sirena also were considerable larger than those of P. emilyae . For these reasons we have decided to maintain the two as separate species, and hope that better material of P. emilyae will be collected in the future in order to confirm or not the original description.
Distribution. Erenna sirena sp. nov. has been collected in the two main MBARI ROV sampling regions, namely Monterey Bay (MB) and the Gulf of California (GOC). All known specimens that have been identified from in situ photographs or collected (emboldened) are listed below.
There are numerous records for Erenna sirena sp. nov. from both the regions of Monterey Bay and the southern part of the Gulf of Mexico. Those from the former region had a mean depth of 2188 ± 175 m, while that for the latter was somewhat shallow at 1849 ± 231.3 m. This difference was also notable for the mean temperatures that were 1.97 ± 0.15°C for the former and 2.46 ± 0.28°C for the latter. However, this may be a reflection of the water depth that, for the latter was c. 3000 m, while for the former it was often> 4000m. Although the known distribution of E. sirena sp. nov. presently is quite restricted it is probable that it has a much broader distribution, but the dearth of studies of the deeper waters of the world's oceans means that it has, to date, been overlooked
Etymology. The specific name sirena is derived from the sirens of Greek mythology, who lured sailors to their death, in reference to the function of the bioluminescent tentilla this species uses to attract its prey.
Genetic results. Molecular sequences for the nuclear 18S and 28S ribosomal RNA genes were largely invariant between the Erenna species, but 16S mtDNA and COI sequences showed a enough variation to potentially aid in identification. There was variation in the 16S and COI between the California and Mexico samples of E. sirena , but otherwise each species was closest to others of the same species, as expected. Sequences were obtained from the following specimens, and have been deposited with GenBank under the accession numbers given below.
The description here of two new Erenna species increases the number of erennids to six. Parerenna emilyae Pugh, 2001 is now the rarest, being known from only a single specimen. However, it is easily distinguished from the Erenna species by the lack of compression of the nectophore in the upper/lower plane, and the indistinctiveness of the ridge system. For the Erenna species, E. insidiator sp. nov. is only known from two specimens, but is clearly distinguishable by the long wing- or fish shaped cnidoband on the tentillum. The maximum size of the nectophore is approximately the same in all species, except Erenna laciniata where they appear to be slightly smaller. However, there are certain characters that should easily distinguish them and these are used in the following key:









| ROV |
Museo Civico di Rovereto |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.