
Conchocarpus cardenasii Londoño-Ech., A.M.
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.601.2.4 |
|
DOI |
https://doi.org/10.5281/zenodo.8134750 |
|
persistent identifier |
https://treatment.plazi.org/id/D512878E-2641-FFE5-2EDD-F897FE97C8EF |
|
treatment provided by |
Plazi |
|
scientific name |
Conchocarpus cardenasii Londoño-Ech., A.M. |
| status |
|
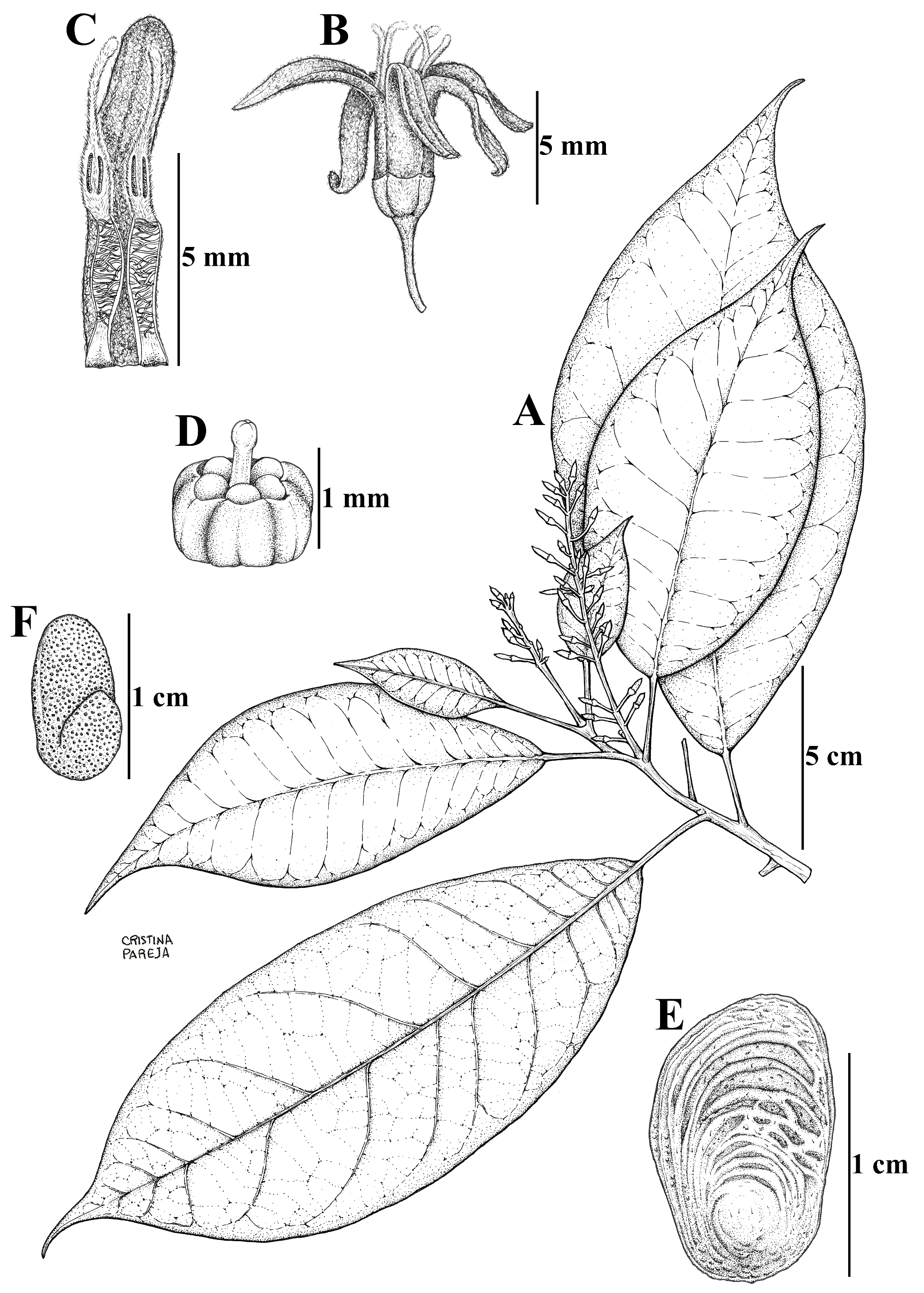
Conchocarpus cardenasii Londoño-Ech., A.M. View in CoL Trujillo & Pérez-Zab., sp. nov. ( Figs. 1 View FIGURE 1 & 2 View FIGURE 2 )
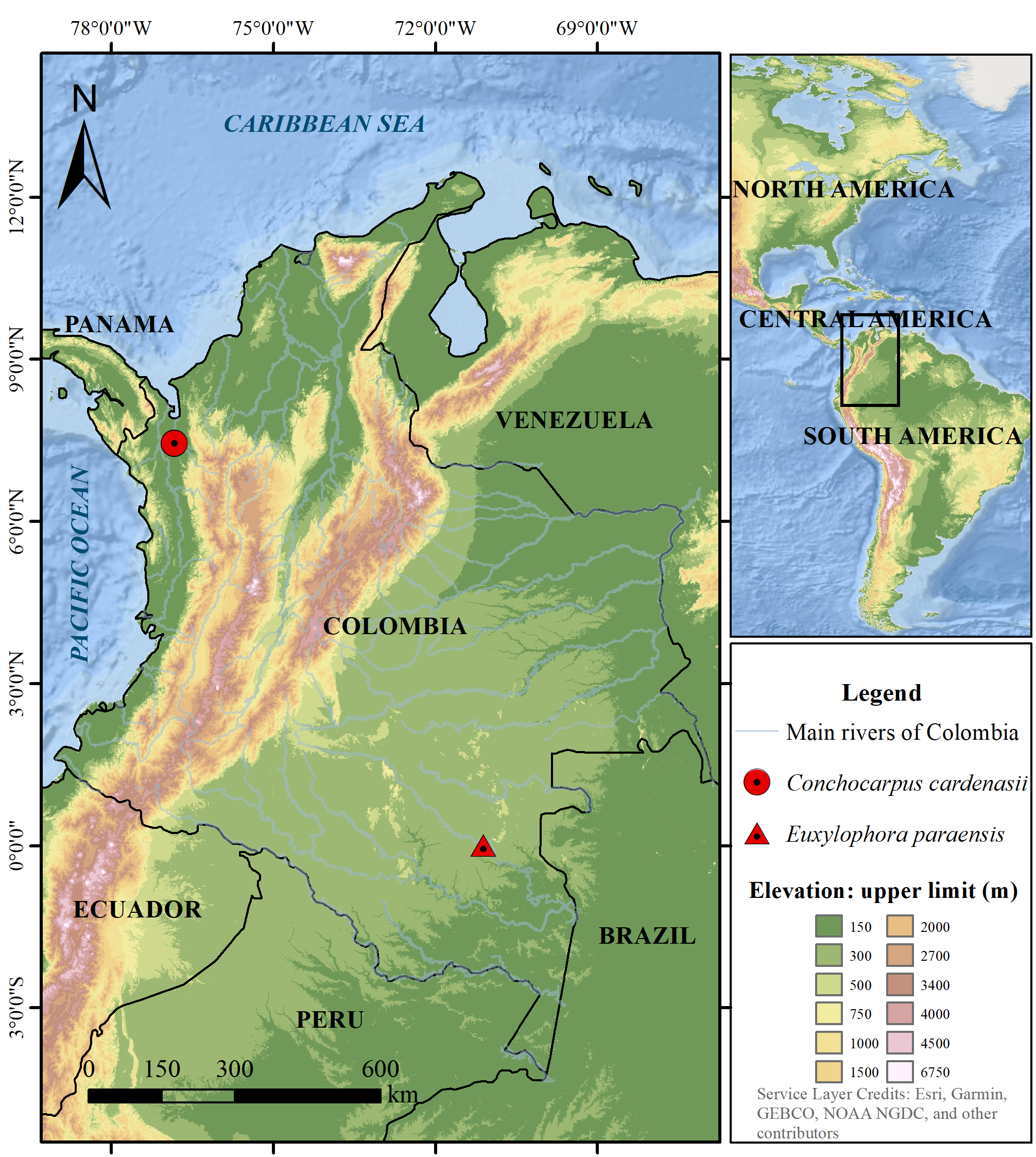
Type:— COLOMBIA. Chocó: Mun. Riosucio, zona de Urabá , Cerros Del Cuchillo , camino La Eugenia (Nova) a la cumbre sureste, 50–300 m, 19 April 1988 (bud & fl), D. Cárdenas 1735 (holotype: JAUM-015131 !) .
Diagnosis:— Conchocarpus cardenasii is morphologically similar to Conchocarpus grandis Kallunki in Kallunki & Pirani (1998: 299), Conchocarpus jirajaranus Kallunki & W. Meier (2019: 196) and in minor degree to Conchocarpus guyanensis ( Pulle 1912: 142) Kallunki & Pirani (1998: 300) . C. cardenasii can be distinguished from the former two species by its shorter petioles, up to 3.3 cm long (versus up to 12 cm long in C. grandis , up to 7.5 cm long in C. jirajaranus ), its longer flowering pedicels 0.7–1 cm long (vs. 0.3–0.6 cm long in C. grandis , ca. 0.2 cm long in C. jirajaranus ), its anthers basally and apically sterile (vs. anthers only apically sterile), its smaller mericarps ca. 1.2 × 0.9–1.1 cm (vs. 2–2.5 × 1.7–2 cm in C. grandis , 1.5–2 × 1.5–1.8 cm in C. jirajaranus ) and its ovary glabrous (only different respect to C. jirajaranus with ovary densely strigulose). From C. guyanensis , the new species can be distinguished by its racemiform thyrses, bearing 15–26 sessile cymules along rachis (vs. long-pedunculate and corymbiform thyrses bearing 3–4 partial inflorescences near apex, each stalked, ca. 2 cm long) and its seeds with plano-convex cotyledons (vs. plicate-conduplicate cotyledons).
Tree 5–10 m tall, branchlets with internodes evident, velvety to densely short-strigose, glabrescent, longitudinally ridged. Leaves 1-foliolate, alternate, the petiole (0.7–) 1.6–3.3 cm long, canaliculate adaxially, short-strigose, glabrescent, ridged longitudinally, transversely striate, swollen at the base and apex; blade (7–)15.5–26 × (3.4–) 5.6– 11.4 cm, elliptic, basally obtuse and cuneate to convex; apically acute and slightly acuminate; sparsely short-strigose on both surfaces, glabrescent; surface pellucid-punctate, the glands visible as dark dots; the margin entire, slightly revolute; secondary veins 12–17 pairs, eucamptodromous becoming brochidodromous distally, raised on both surfaces, more distinct abaxially, intersecondary veins present; tertiary veins percurrent, raised on both surfaces, more distinct abaxially. Inflorescence a racemiform thyrse, 4.7–19.1 cm long (including peduncle), short-strigose and puberulous, slightly ridged longitudinally, apparently supra-axillary (originating from a terminal meristem but reoriented laterally by the sympodial growth), the axillar vegetative bud persisting below; the peduncle 0.9–4.8 cm long, the flowering portion 2.5–14.3 cm long, bearing 15–26 flowering nodes, each a dense and sessile cymule, 1–3-flowered, the axes indiscernible; the primary bracts 1–2 × 1 mm, ovate, subtending each cymule, densely short-strigose on abaxial surface and on adaxial surface only towards the margin (glabrous toward the middle portion); the secondary bracts 0.2–0.5 × 0.2–0.5 mm, ovate, subtending some pedicels, densely short-strigose on abaxial surface and on adaxial surface only towards the margin (glabrous towards the middle portion), pedicels 0.7–1 cm long, short-strigose and puberulous, glabrescent. Flowers 5(–6)-merous, calyx cupular, 1.5–1.9 mm high, 5-dentate, the teeth glandular ca. 0.2 × 0.2 mm, densely to sparsely short-strigose abaxially, glabrous adaxially; corolla actinomorphic, cochlear, white, short-tubular by capillinection, the tube ca. 4.5 mm high; petals 7–9.6 × 1–1.6 mm, oblong, apically obtuse and rounded to slightly acuminate, coherent into a tube among them, tomentose abaxially, adaxially glabrous at the base and near to the portion adnate to the filaments, the rest of surface tomentose, on the mouth of tube mixed with little short-villous indumentum, pellucid-punctate more noticeably abaxially; androecium 5(–6)-merous, all stamens fertile, filaments 3–3.7 × 0.6–0.8 mm, oblong to narrowly oblanceolate, coherent among them by capillinection, adherent to the corolla at the base and adnate to it only by a small dorsal portion at the tube throat (perhaps postgenital fusion, see discussion), abaxially tomentose, glabrous near to the portion adnate to the corolla, adaxially glabrous at the base, the rest of surface tomentose, mixed with short-villous indumentum at the throat; anthers 3.2–4 × 0.5–0.7 mm, narrowly lanceolate, broadly attached to the filaments, sterile at the base and at the apex, eglandular at the connective, not exserted along their whole length from the corolla tube; thecae with a basal sterile portion of 0.2–0.4 mm long, a middle fertile portion of 1.5–2.1 mm long, abaxially sparsely short-villous, adaxially short-villous; the apical sterile portion 1.3–1.6 mm long, flattened, curved in anthesis, abaxially sparsely tomentose, adaxially sparsely tomentose mixed with little short-villous indumentum; disc ca. 0.7 mm high, glabrous, slightly shorter than the ovary, margin sinuate; gynoecium 5-merous, apocarpic, glabrous, not exserted from the throat at anthesis, ovary 0.8–0.9 mm high, ca. 0.8 mm diam., umbilicate, style ca. 0.3 mm long (measured over ovary apex), stigma 0.2–0.3 mm long, capitate. Fruits composed by a single mericarp, ca. 1.2 × 0.9–1.1 cm, semi-orbicular laterally, ventrally straight, dorsally rounded, very sparsely short-strigose, pellucid-punctate, transversely ridged. Seed 1 per carpel, ca. 1 × 0.8 cm, glabrous, slightly rugulose, testa papery, embryo straight, cotyledons plano-convex, thick.
Distribution and habitat:— Conchocarpus cardenasii is endemic to Colombia. It has been collected only in a hill named as “Cerro del Cuchillo” on the Riosucio municipality of the Chocó department ( Fig. 3 View FIGURE 3 ). The Cerro del Cuchillo is a small and isolated hill on the Pacific Biogeographical Region (sensu Bernal 2016) in a range of elevation of 20–450 m, with annual precipitation between 4.000 to 8.000 mm ( Cárdenas-López 2003). This region is considered under the Equatorial Rainforest climate (Af climate type sensu Kottek et al. 2006).
Phenology:— Herbarium vouchers indicate blooming time in April and fruiting time in June.
Etymology:— The epithet honors to Dairon Cárdenas, a noteworthy Colombian botanist who dedicated most of his career to improving knowledge of the Colombian Amazonian flora. The only known specimens for the new species were collected by Dairon early in his career. This is a modest acknowledgment to Dairon, for his assiduous work that rendered so many contributions to the knowledge of the flora, also to his kindness and friendship with the authors when they visited the COAH herbarium.
Preliminary conservation status:— Conchocarpus cardenasii is only known by a subpopulation with AOO= 4 km 2. The forest that inhabits is a small fragment <70 km 2 immersed in a matrix of severe deforestation with two important human settlements less than 20 km away (i.e., the unique known population is severe fragmented and under continuous decrease of habitat quality). Although it can be classified as “Critically endangered” according to the criteria B: B2ab(iii) of IUCN (2012), the Cerro del Cuchillo is currently a protected area, which decreases the effect from any threat. It should be rather considered as “Endangered” species, due to its small and restricted geographical distribution, surrounded by livestock pastures with null ecological connectivity with near forests.
Additional specimens examined (paratype):— COLOMBIA. Chocó: Mun. Riosucio, zona de Urabá , Cerros Del Cuchillo , camino de Cidon a la Cumbre sureste, 50–100 m, 23 June 1988 (fr), D. Cárdenas 2090 ( JAUM-015132 !) .
Notes:— Conchocarpus cardenasii has a close resemblance to C. grandis and C. jirajaranus . These three species share the 1-foliolate leaves, the racemiform thyrses with partial inflorescences sessile (only stalked at the lowermost ones in C. grandis ), the cupular calyx, the actinomorphic corolla (not verified for C. grandis ), the all-fertile stamens, the lanceolate and apically sterile anthers, the umbilicate ovary (not verified for C. jirajaranus ) and the seeds with plano-convex cotyledons. On the other hand, C. guyanensis shares with the new species the unique in the genus presence of anthers with thecae basally sterile, but can be easily differentiated from it and the other two species above mentioned by its long-pedunculate and corymbiform thyrses and seeds with plicate-conduplicate cotyledons. Other features are relatively similar among the four species discussed ( Kallunki & Pirani 1998, Kallunki & Meier 2019), see Table 1 View TABLE 1 .
The filaments of Conchocarpus cardenasii are here described as adnate to the corolla only by a small dorsal portion at the tube throat, because they are strongly united and hardly detachable without being torn apart. El Ottra et al. (2019) recently reported a kind of postgenital floral fusion in Galipeinae consisting in the union by papillae or cuticular projections; whether this is the situation responsible for the structure here observed in C. cardenasii remains a matter depending on further anatomical studies.
The cotyledons plano-convex in Conchocarpus cardenasii , C. grandis and C. jirajaranus are morphologically atypical for Conchocarpus sensu stricto, and its taxonomic relevance could be tested in further phylogenetic studies. Similarly, other species currently under Conchocarpus have uncommon features that could support new classification proposals. For example, Conchocarpus nicaraguensis [ Standley & Williams (1953: 206–207)] Kallunki & Pirani (1998: 314) and Conchocarpus ucayalinus [ Huber (1906: 573–574)] Kallunki & Pirani (1998: 323) from northwestern South America, Central America and western Amazonia, resemble at first glance species of Dryades because their calyx lobes imbricate and overlapping even after anthesis. However, not all diagnostic features of Dryades are present in C. nicaraguensis and C. ucayalinus and these species have not been included in phylogenetic analyses. Added to the morphological relevance of the cotyledons in the classification of Conchocarpus sensu lato, Groppo et al. (2017) found Conchocarpus toxicarius [Spruce ex Engler (1874: 114–115)] Kallunki & Pirani (1998: 322), phylogenetically segregated from Conchocarpus s.s. and Dryades (this genus not segregated yet in Groppo et al. 2017), besides noting it under the character state “cotyledons plane or plane-convex” (see Table 1 View TABLE 1 ), which suggest that this species perhaps might be recognized under another genus when a more complete sampling becomes available (Groppo, pers. comm. 2018).
Interestingly, although Conchocarpus toxicarius can be distinguished by its leaves 5–7-foliolate from C. cardenasii , C. grandis and C. jirajaranus , all four species share their androecium with all stamens fertile, lanceolate anthers and in a sensu lato, their actinomorphic corolla, umbilicate ovary and plano-convex cotyledons (see Table 1 View TABLE 1 ). Remarkably, one of the two syntypes of C. toxicarius belongs to Conchocarpus heterophyllus [A. Saint-Hilaire (1823: 131)] Kallunki & Pirani (1998: 300), an inconsistence solved by Kallunki & Pirani (1998); thus, although the protologue of C. toxicarius indicated only three fertile stamens, Kallunki & Pirani (1998) indicated that it have five stamens fertile, which is according to the original illustration at the complete flower drawings [see Engler (1874: t.24, fig. 1) and the lectotype Spruce 2171 (K barcode 000531071 [digital image!]); note the original illustration at stamens details drawings showing those lanceolate anthers of C. toxicarius as staminodes and probably, those of C. heterophyllus as fertile stamens. Currently, the recognition of other segregate groups from Conchocarpus is not definitive considering the lack of sampling species with deviating morphological combinations in molecular analyses (e.g., actinomorphic corolla, five stamens fertile, lanceolate anthers and plano-convex cotyledons).
Despite of the fact that Conchocarpus cardenasii could undergo nomenclatural and taxonomic changes if the splitting of Conchocarpus is supported in future phylogenetic studies, its morphological features fits into the current circumscription of the genus. Considering the remarkably presence of cotyledons plano-convex and anthers basally sterile, C. cardenasii may be a critical species to be included in future morphological and DNA systematics studies of the genus. Finally, given its condition of being a restricted endemic and endangered species, this formal taxonomic recognition is fundamental for supporting current and future conservation actions at the unique Cerros del Cuchillo remnant forests.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Zanthoxyloideae |
|
Genus |