
Bythotrephes transcaucasicus Behning, 1941
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4138.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:724ABC9B-9548-4E3A-A667-274F10A19C8E |
|
DOI |
https://doi.org/10.5281/zenodo.6085647 |
|
persistent identifier |
https://treatment.plazi.org/id/D5745718-9B40-FFF0-FF03-FBABFDE8FEF6 |
|
treatment provided by |
Plazi |
|
scientific name |
Bythotrephes transcaucasicus Behning, 1941 |
| status |
|
Bythotrephes transcaucasicus Behning, 1941 View in CoL
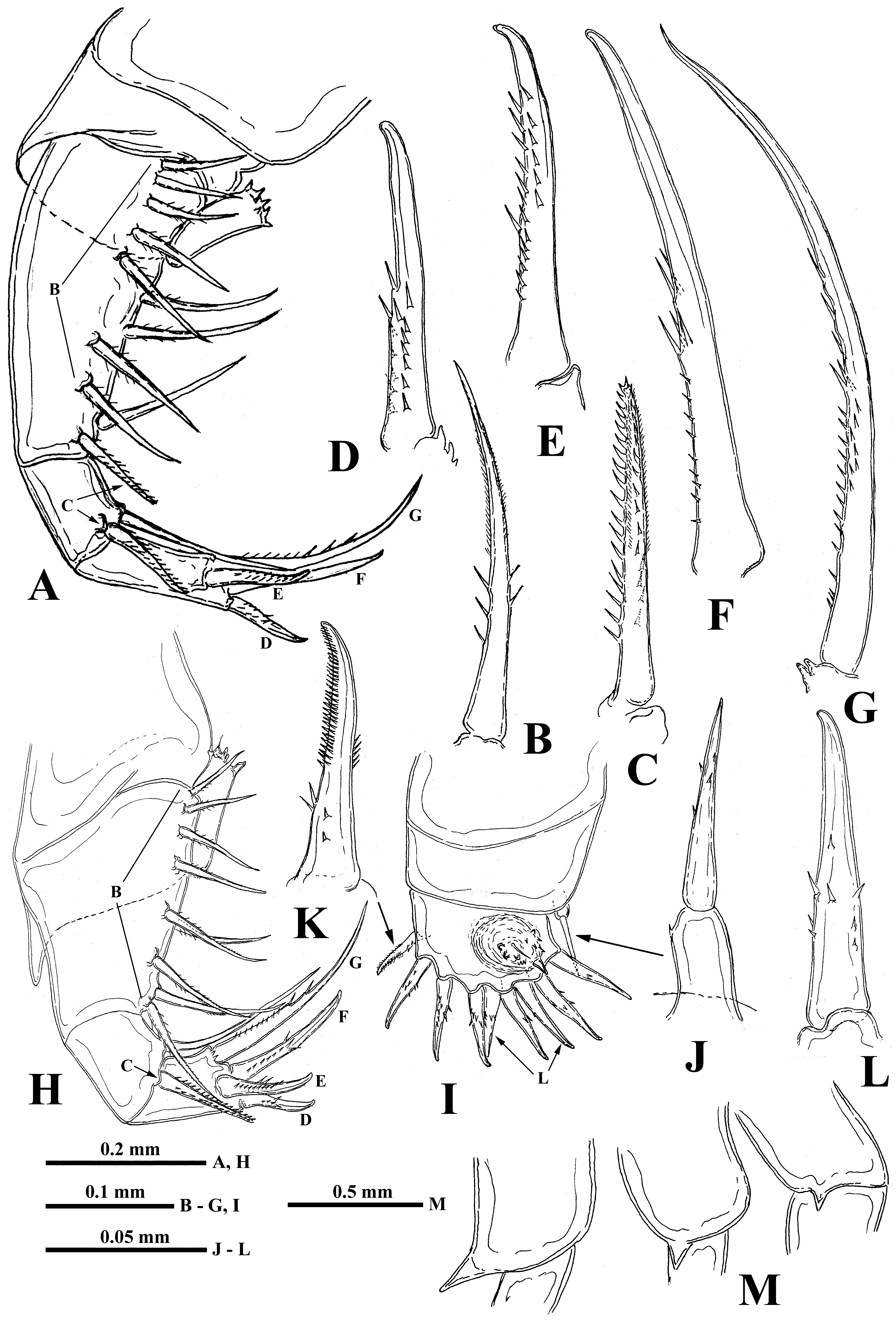
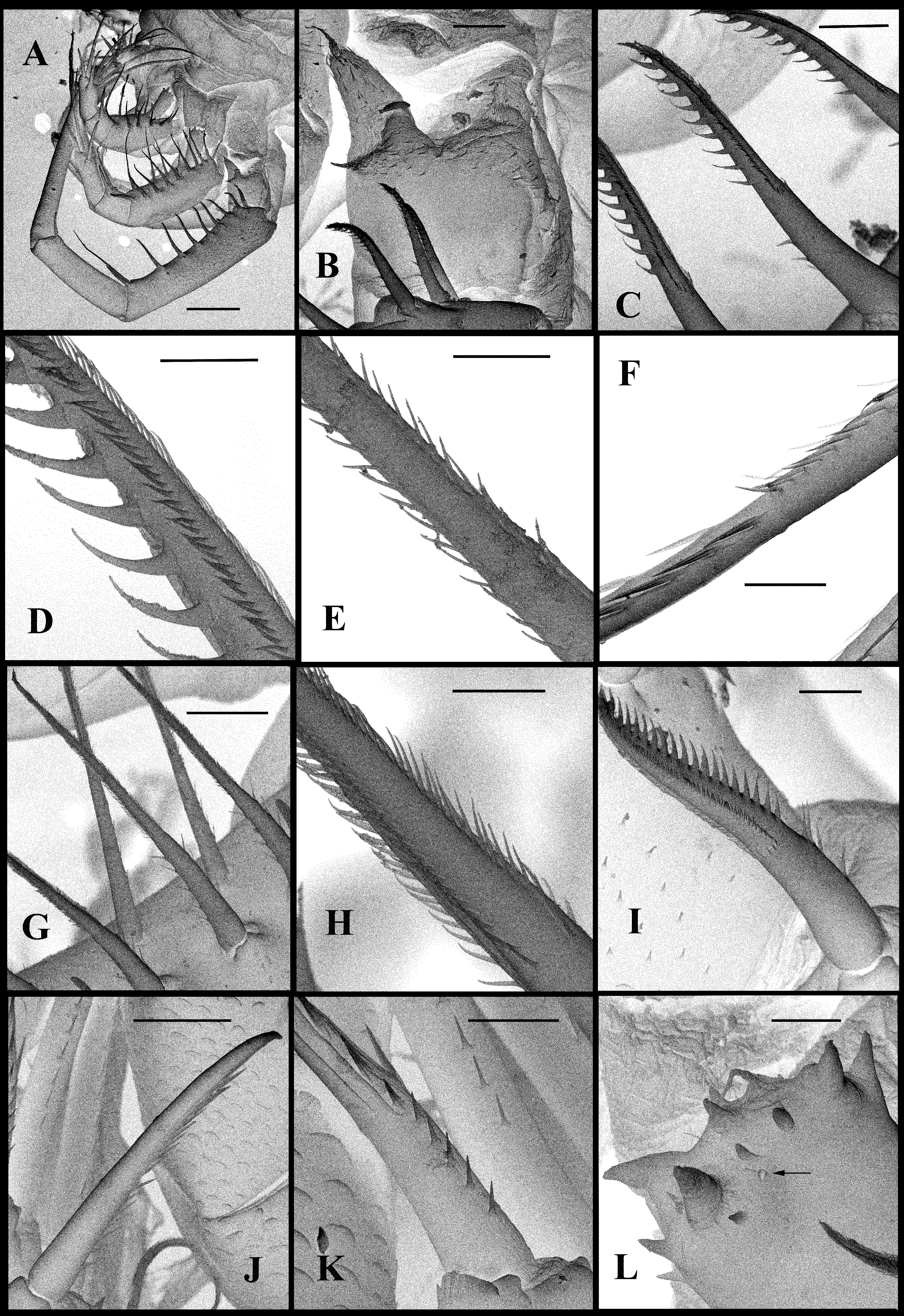
( Figs. 7 View FIGURE 7 , 8 View FIGURE 8 )
Bythotrephes longimanus transcaucasicus Behning, 1941 View in CoL : Behning 1941: 355 –356; Litvinchuk 2001: 129 Bythotrephes transcaucasicus Behning, 1941 View in CoL : Litvinchuk 2002: 129, Fig. 39, 41B; 2007: 192–193, Fig. 3 View FIGURE 3 B; Litvinchuk & Litvinchuk 2016: 67 –73, Fig. 9 View FIGURE 9 .
Type material. Because Behning (1941) analyzed specimens from Lake Chaldyr, they must be considered specimens of the type series of the taxon ( ICZN,), syntypes ( ICZN, 72.4.1., 72.5., 73.2.).
In the course of the present taxonomic revision, a lectotype and paralectotypes were designated ( ICZN, 74.1.).
Lectotype. A female with body length 3.73 mm from Lake Chaldyr ( Turkey) collected by A. Brandt in 1879 ( ZIN, № 55175).
Paralectotypes. All other specimens either on slides or in liquid samples are considered (ZIN, slides № 55165–55170, samples (subsamples) № 55171–55174).
Material examined. Turkey: 1) sample ( ZIN, N 1635, two subsamples in two vessels) labeled “ Bythotrephes longimanus Leydig, Kuchik (Chaldyr) , A. Brandt, 1879”, 2 ad (paralectotypes); 2) sample ( ZIN, N 1636, two subsamples in two vessels) labeled “ Bythotrephes longimanus Leydig, Lake Chaldyr , on the surface, A. Brandt, 1879”, 20 ad (paralectotypes, lectotype was selected from this sample, see below); 3) six slides ( ZIN, without numbers) labeled “ Bythotrephes, Chaldyr ” or “ Bythotrephes longimanus Leyd, Lake Chaldyr, Brandt, 1879 ”, 11 ad (paralectotypes).
Data on body and body parts measurements of specimens of the species are presented in Table 1.
Description. Parthenogenetic female. General body appearance and segmentation as in previous species ( Fig. 7 View FIGURE 7 A).
Head comparatively large (34.0–38.5 % of body length) and subdivided into two parts: rounded anterior part mostly filled by large compound eye and posterior part bearing dorsally a large saddle-shaped neck organ, swimming antennae and mouth parts. Large pigment spot occupies about one-third or more of the eye’s volume; ocellus (naupliar eye) is absent.
Antennules small and situated on the ventral side of the anterior part of head beneath the eye. They are bulbous ( Fig. 7 View FIGURE 7 B) and sit on the joined basis, slightly splitted anteriorly. Terminally, they bear five regular aesthetascs in two groups, in three and two in each one, one shorter and thinner aesthetasc-like structure situated in a group with two regular aesthetascs, and having slightly widened apical end with a dark granular structure inside.
Swimming antennae. Comparatively long, with elongated cylindrical basipodite ( Fig. 7 View FIGURE 7 A). Of two antennal branches, the lower three-segmented one (endopodite) ( Fig. 7 View FIGURE 7 D) sitting on the apical basipodital prominence, is slightly longer than the upper branch. The upper branch ( Fig. 7 View FIGURE 7 C) is four-segmented and lower branch is threesegmented. Proximalmost segment of the upper branch is rudimentary and clearly visible only externally; all other segments of both branches are much more developed. Small spine with a row of the neighboring minute prominences on the end of second segment of upper antennal branch; similar apical prominences and spines on the distal segments of both branches ( Figs. 7 View FIGURE 7 E, 7F). Small proximalmost segment of upper branch lacks setae, while other segments possess two-segmented swimming setae of more or less similar size except distal of them which are shorter. All setae bilaterally armed with rows of uniform thin setulae. General formula of antennal setae: 0–1–2–5 / 1–1–5.
Mouth parts are generally similar to those of previous species. They were not investigated in detail because of the presence of only comparatively few type specimens available.
Carapace. It looks like a bag-like structure, strongly modified into closed brood pouch ( Fig. 7 View FIGURE 7 A) connected in its wide base with dorsal side of thorax.
Thoracic limbs. Four pairs of strongly chitinized, stenopodous limbs are densely situated along the muscular ventral side of thorax and directed antero-ventrally ( Figs. 7 View FIGURE 7 A). All of them have complex and variously setaceous armament along their inner side. Limbs of the three anterior pairs are five-segmented and those of the fourth pair are three-segmented. Protopodites of all of them, covered by comparatively softer cuticle, are inconspicuously delimited into two parts (segments), coxa and basis; the endopodites of limbs of the three anterior pairs are composed of three well developed segments; those ones of the fourth pair are unisegmented ( Figs. 7 View FIGURE 7 A, 7G, 8C, 8E, 8G).
First pair of limbs (tl I) are especially long and strong; their length shorter than body length (68.0–77.9 % of body length) ( Figs. 7 View FIGURE 7 G). Terminally, the inner side of their protopodite bears a small triangular lobe, the pseudognathobasic process (see explanation of the term in Korovchinsky (2015)), armed laterally and distally with two outgrowths with apical setae and numerous spinulae ( Fig. 7 View FIGURE 7 G, 7H). The external part of the protopodite is longer than the internal one and bears apically a small conical outgrowth ( Fig. 8 View FIGURE 8 A). The first segment of the endopodite is long (21.5–24.6 % of body length) and bears 6–7 anterior lateral setae (their number on limbs of one individual can vary) with a posterior row of incurved spines, and anterior and lateral rows of fine setulae (as in B. arcticus : Figs. 3 View FIGURE 3 C, 3D). Distally, this segment bears shorter anterior seta of the same type and a long posterior finely setulated seta ( Figs. 7 View FIGURE 7 I, 7J). The second segment of the endopodite is conspicuously shorter (11.6–15.6 % of body length) and bears only two apical setae similar to those on the end of previous segment but shorter ( Figs. 7 View FIGURE 7 K, 7L). The terminal third segment of the endopodite is also long (18.8–23.2 % of body length), almost equal to or slightly shorter than the length of the first segment, and always bearing apically four long roughly spinulated setae, two of them terminally and two subterminally ( Fig. 7 View FIGURE 7 G). Basally, these setae are armed by a row of smaller spines, while distally, by larger lanceolate spines situated in two rows and directed terminally (as in B. arcticus : Fig. 3 View FIGURE 3 F).
Second pair of limbs (tl II) are considerably shorter, their protopodite, again externally, is conspicuously longer and bears apically a conical outgrowth ( Figs. 8 View FIGURE 8 B-right, 8C). The first, basal segment of the endopodite bears a row of 6–8 (mostly 6–7) rather long anterior lateral setae (their number also can vary in one individual) ( Figs. 8 View FIGURE 8 C, as in B. arcticus : 2B, 3G, 3H). There are 2–3 posterior lateral seta of the same type on this segment. The terminal setae of the segment are different; the anterior one is shorter and roughly armored (as in B. arcticus : Figs. 2 View FIGURE 2 C, 3I), while posterior one is longer and finely setulated. Internally, this segment bears a stout cylindrical pseudognathobasic process, possessing some prominences of different size and one small, thin seta apically ( Figs. 8 View FIGURE 8 C, 8D). The second segment of the endopodite is short with only two setae, the anteriormost of which is similar to the distal anterior one of the previous segment, while the posterior seta is longer and finely setulated. The distal segment of the endopodite bears four setae, two terminal and two subterminal ones ( Fig. 8 View FIGURE 8 C). Of the latter, the anterior seta is comparatively short, thick and armed with a number of thin lateral denticles, while its distal end is naked and slightly hooked apically (as in B. arcticus : Fig. 2 View FIGURE 2 E, 3J). Its neighboring posterior subterminal seta is considerably longer and similar to the subterminal and terminal setae of tl I, having similar spine armament and sharp apex (as in B. arcticus : Fig. 2 View FIGURE 2 G). The anterior terminal seta is thick and comparatively short with longitudinal ribs, few thin lateral denticles, and slightly hooked apical end (as in B. arcticus : Figs. 2 View FIGURE 2 D, 3K). The posterior terminal seta is similar to the neighboring anterior one but longer; it bears few lateral denticles and has a slightly hooked apical end (as in B. arcticus : Fig. 2 View FIGURE 2 F).
Third pair of limbs (tl III) are generally similar to those of the previous ones, differing in some details. The external outgrowth of their protopodite is conspicuously larger ( Fig. 8 View FIGURE 8 B– left), and lateral anterior and posterior setae of the first segment of the endopodite are fewer (5–6 and 1–2, respectively) ( Fig. 8 View FIGURE 8 E). Distal setae of the segment are similar to other ones. The pseudognathobasic process is also similar to that of tl II ( Figs. 8 View FIGURE 8 E, 8F). About setae of the second segment, the anterior one is similar to the respective one of tl II (as in B. arcticus : Fig. 4 View FIGURE 4 A). The terminal and subterminal setae of third segment are similar to those of tl II but slightly shorter and bearing fewer denticles ( Fig. 8 View FIGURE 8 E, as in B. arcticus : 4B, 4C).
Fourth pair of limbs (tl IV) are considerably reduced; their protopodite bears a slightly spinulated seta situated on a short cylindrical base ( Fig. 8 View FIGURE 8 G, as in B. arcticus : 2J). The only segment of the endopodite has two rows of comparatively short spine-like setae. The external row (group) always consists of two setae, and the internal row consists of 6–7 setae, which differ in their appearance and armament (as in B. arcticus : Figs. 2 View FIGURE 2 K, 2L, 4E, 4F). Almost the whole internal part of the terminal segment is occupied by the reduced pseudognathobasic process armed by some denticles and a thin seta ( Figs. 8 View FIGURE 8 G, 8H).
Abdomen (metasome) ( Fig. 7 View FIGURE 7 A) is often deformed. It is inconspicuously delimited in two segments, a short proximal and a long distal ones with or without a prominent fold situated more or less in the middle of the dorsal side.
“ Postabdomen” is comparatively small, actually consisting of two parts, the last small abdominal segment and the postabdomen per se (see Korovchinsky (2015)) ( Figs. 7 View FIGURE 7 A, 8L). The postabdominal claws are large (6.0–11.1 % of body length), always curved backwards ( Figs. 7 View FIGURE 7 A, 7M, 8I –K).
Caudal process is directly connected with the postabdomen and proceeds as a very long, proximally rather thick and curved structure, then a straight spine-like form ( Fig. 7 View FIGURE 7 A, 7N), variable in length (127.1–157.9 % of body length), exceeding the body length by 1.2–1.6 times. Basally, the caudal process bears one, rarely two pairs of claws similar to those on the postabdomen but usually smaller (e.g., proximal claws reach 5.0–9.6 % of body length), and apically two minute setae arise from a common base ( Fig. 7 View FIGURE 7 O) (they were observed only in few specimens). There is usually only one pair of claws on the process; two pairs of such claws were observed in only one female of 29 investigated. Pairs of claws sit closely (e.g., distance between postabdominal claws and proximal claws of caudal process (interclaw distance) constitute 7.6–9.9 % of body length). Between these pairs of claws, the thickness of the structure ( Figs. 7 View FIGURE 7 M, 8I –K), reaches 4.4–7.2 % of body length. Borders separating old molted integuments of caudal process with claws either are barely visible or clearly visible.
Body length. 2.3–4.2 mm.
Males and gamogenetic females are not known.
Remarks. The level of morphological variability of population of the species seems rather low, the comparative length of claws is most variable (see Table 1).
Behning (1941) recorded B. longimanus transcaucasicus from three Transcaucasian lakes, Chaldyr ( Turkey), Tabatsquri ( Georgia), and Arpa lake (Arpi-lich lake) ( Armenia). He had only eight specimens at his disposal without indication of their locality, probably not knowing about the presence of more extensive material in ZIN collection.
In comparison with Behning’s (1941), Litvinchuk’s (2002, 2007), and Litvinchuk & Litvinchuk (2016) data, specimens studied in the present work have larger body size (up to 4.2 mm) probably as a consequence of analysis of larger number of specimens. The latter authors correctly mentioned shorter claws of B. transcaucasicus in comparison with B. arcticus .
Differential diagnosis. The present species seems closer to B. arcticus . It differs from the latter in few inconspicuous features: presence of comparatively shorter claws, smaller distance between claws which are closer situated, and smaller thickness of caudal process between postabdominal claws and proximal claws of caudal process (differences in these characters between representatives of B. transcaucasicus and sets of B. arcticus specimens ( Table 1) in pairwise comparison were statistically significant: p <0.01).
The smaller body size and presence of predominantly two pairs of claws in the specimens of B. transcaucasicus , one on the postabdomen and other on the proximal part of the caudal process, can be specified as the additional differences. Of the studied specimens, three pairs of claws were observed in only one individual of 29 studied ones, while in B. arcticus specimens with three pairs of claws often were present in all samples. However, these features should be examined further because only comparatively limited material of B. transcaucasicus has been investigated.



| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SuperOrder |
Cladocera |
|
Order |
|
|
Family |
|
|
Genus |
Bythotrephes transcaucasicus Behning, 1941
| Korovchinsky, Nikolai M. 2016 |
Bythotrephes longimanus transcaucasicus
| Litvinchuk 2016: 67 |
| Litvinchuk 2002: 129 |
| Litvinchuk 2001: 129 |
| Behning 1941: 355 |