
Egle concomitans ( Pandellé, 1900 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2043.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/D73DC225-6D57-902B-FF73-4A3CFEBC9D6D |
|
treatment provided by |
Felipe |
|
scientific name |
Egle concomitans ( Pandellé, 1900 ) |
| status |
|
1. Egle concomitans ( Pandellé, 1900) View in CoL
Figs. 27 View FIGURES 27–30 , 31–38 View FIGURES 31–38 , 52–54 View FIGURES 52–56 .
Anthomyia (Chortophila) concomitans Pandellé, 1900: 251 View in CoL , 287.
Chortophila cacuminata Villeneuve, 1923: 95 View in CoL . Synonymized by Hennig (1972: 429).
Hylemyia hinei Malloch, 1920b: 278 View in CoL , figs. 5, 26, 33. Synonymized by Darvas (2001: 394).
Hylemyia ithacensis Huckett, 1924: 28 View in CoL , figs. 7, 63, 112. Synonymized with Hylemyia hinei Malloch View in CoL by Huckett (1965: 66).
Chortophila concomitans (Pandellé) ; Séguy 1923: 123, fig. 187.
Chortophila (Nudaria) concomitans (Pandellé) ; Karl 1928: 173.
Pegohylemyia concomitans (Pandellé) : Hennig 1967b: 179, text fig. 164.
Lasiomma concomitans (Pandellé) ; Hennig 1972: 429, text fig. 380, plate figs. 530, 607, 675; Ackland & Pont 1977:
444; Rozkošný & Barták 1984: 465; Fan et al. 1988: 63, figs. 128–130; Hua 2006: 100.
Egle concomitans (Pandellé) View in CoL ; Michelsen 1988: 277; Barták et al. 1989: 316; Barták et al. 1990: 441, 446; Dely- Draskovits 1993: 51; Wei et al. 1998: 660, fig. 1472; Meixnerová & Rozkošný 1999: 383; Teschner 1999: 131; Darvas 2001: 394; Darvas et al. 2001: 309; Michelsen & Barták 2001: 457; Ackland & Merz 2003: 210; Griffiths 2003: 2293, figs. 2585–2593; Michelsen 2004; Hua 2006: 98; Komzáková 2006.
For further references, see Griffiths (2003).
Description. Medium-sized, wing length 4.0–5.4mm.
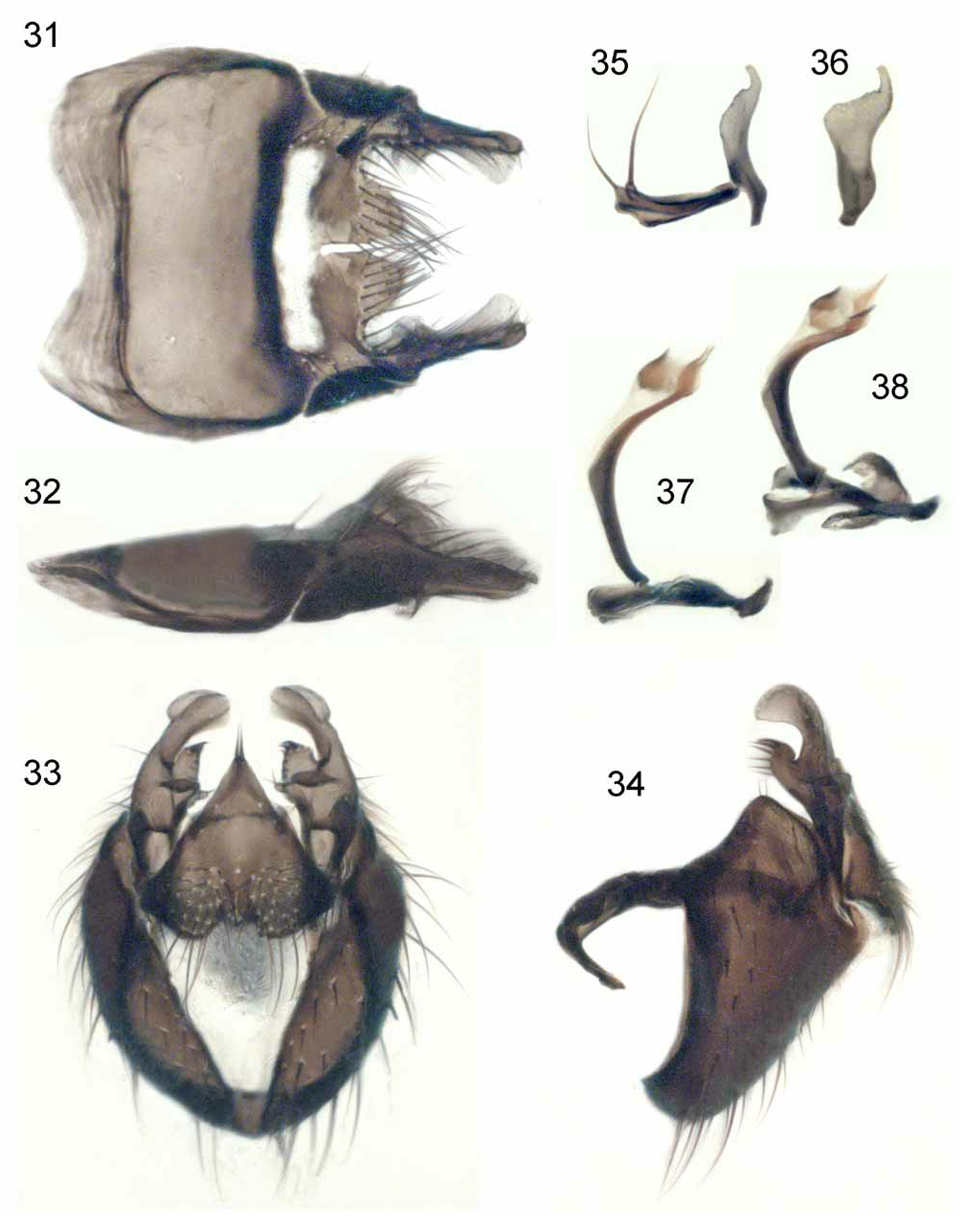
Male. Body covered in light greyish dusting; mesonotum in rear aspects with three broad brownish black stripes, lateral ones widely interrupted in front of suture; abdomen with mid-dorsal dark stripe and dark bands basally on tergites. Chaetotaxy of head, body and legs unremarkable, not long and dense; main setae well differentiated from setulae. Head ( Fig. 27 View FIGURES 27–30 ) much higher than long. Frons very narrow on upper part, usually less than 0.5 times diameter of anterior ocellus and with contiguous parafrontalia. Pair of interfrontal setulae in middle of frons standing immediately above series of frontal setae; orbital setulae absent. Parafacial in broadest aspect parallel-sided, narrower than postpedicel. Face with weak interantennal keel, weakly projected lower margin in profile lying slightly behind fronto-parafacial angle. Gena short, relatively broad, with setae in several irregular rows. Antennal postpedicel ( Fig. 21 View FIGURES 21–26 ) comparatively long. Proboscis short, comparatively weak; haustellum and palp both ca. 0.4 times length of fore tibia; prementum with extensive grey dusting. Proepisternals 2; proepimerals 6–8. Prealar seta about same length as posterior notopleural seta. Lower calypter nearly as big as upper calypter. Fore tibia with 0 ad-, 0 pd- and 1–3 short pv-setae; mid tibia with 1 ad-, 1 pd- and 2–3 p-setae; hind femur with a row of pv-setae interrupted basally and subapically; hind tibia with 4–6 av-setae and a more or less complete row of short pv-setae. Terminalia ( Figs. 31–38 View FIGURES 31–38 ): cerci, surstyli and gonites of distinctive shape.
Female. Differences from male (apart from usual sexual ones): Body lighter greyish dusted, finely tinged with brown on vertex and mesonotum; pattern of dark brown striping on mesonotum very faint; grey dusting on abdomen with strong dark reflections obscuring narrow mid-dorsal dark stripe. Genal setae fewer, anteriorly in single row. Proepimerals 3–6. Fore tibia with 0–1 ad-, 0–1 pd- and 1–2 pv-setae. Mid tibia with 1–2 ad-, 1–2 pd- and 2–3 p-setae. Hind femur with only single pv-seta apically; hind tibia with 2–4 av-setae. Oviscapt ( Figs. 52–54 View FIGURES 52–56 ): Sternites VI and VII complete but narrow, each with pair of hind marginal setae; sternite VII pieces large, each with two setae at hind margin; epiproct with several setulae in addition to pair of setae; cerci rather short, broadly rounded on apical part. Three similar, moderate-sized spermathecae with cross-striae.
Material examined. CZECH REPUBLIC [ ZMUC]: Central Bohemia: Praha-Troja, along river, 200m, 7 males 28.iv.1991, 1 male, 2 females 16.v.1985 (M. Barták); Praha-Hološovice, river bank, 200m, 3 males 16.v.1984 (M. Barták); Lysá nad Labem env., car net, 1 male 2.v.1992 (M. Barták) . Ústi nad Labem: Bílina- Vĕtrák , dump restoration, 200m, 1 male 23–30.iv.1997 (M. Barták) . South Moravia: Pálava Reserve , lowland wood, 160m, 13 males 9.v.1997 (M. Barták) . FINLAND [ FMNH]: South Karelia: Parikkala , 1 male 19.v.1957 (L. Tiensuu) . SLOVAKIA [ ZMUC]: Nitra : Štúrovo- 1km E, lowland wood, 110m, 1 male 24.iv.1986 (M. Barták). Also seen from CANADA: Quebec and USA: Alaska, Wisconsin, New York .
Biology. In North America, according to Griffiths (2003), adults of Egle concomitans been found in spring on flowering willow together with other species of Egle , but they also regularly visit other flowers later in the season and have even been taken on moose dung and baits of liver and carrion. Unfortunately, the host plant(s) and habits of the expectedly seed-feeding larvae remains unknown. As the adults evidently interact less intimately with the host plants of their larvae than seen in other species of Egle , I find it most likely that larval development takes place in female catkins of poplar and aspen ( Populus spp. ), as these are windpollinated and probably only attractive for egg-laying purposes. The pronounced continental distribution of E. concomitans in Europe may suggest a primary association with native species of poplar, e.g., Black poplar ( Populus nigra L.) or White poplar ( P. alba L.), growing preferably in flood plain forests.
Distribution. A widespread Holarctic species reaching the northern border of Oriental Region. Apparently rare in Europe with a mainly continental and eastern occurrence, but possibly under-collected because exclusively an early-season species. Described from ‘Prusse orient.’, which refers to the present Russian enclave Kaliningrad Oblast, and from France ( Pandellé 1900, Villeneuve 1923). Subsequently recorded from Germany ( Hennig 1972), Czech Republic ( Rozkošný & Barták 1984, Barták et al. 1989), Slovakia ( Barták et al. 1990), Hungary ( Darvas et al. 2001), Switzerland ( Ackland & Merz 2003) and Finland ( Michelsen 2004). Further Palaearctic records are from Mongolia (D.M. Ackland in litt.) and China: Heilongjiang, Liaoning, Beijing, Shanxi and Qinghai ( Fan et al. 1988, Wei et al. 1998, Hua 2006). Reaching the Oriental Region in northern Myanmar: Kachin ( Ackland & Pont 1977; Griffiths 2003). Widespread in boreal and floodplain forests of northern North America ( Griffiths 2003).
Relationships. The short and deep head profile and short proboscis ( Fig. 27 View FIGURES 27–30 ) make Egle concomitans very different from other species of Egle . Still, the detailed agreement in structural details of the male terminalia suggests that it is more closely related to the E. longirostris species group than to the remaining species of Egle . This implies that a slender and extended proboscis has been attained through homoplasy in the E. longirostris group and in the remaining species of Egle . The extended but very shallow pregonite with two strong setae found in E. concomitans is strikingly different from the large, bilobed and abundantly setated postgonite found in the three described species of the E. longirostris group. Both conditions appear apomorphic in Egle , but two undescribed species of the Egle concomitans section from Pakistan (D.M. Ackland, in litt.) combine a more plesiomorphic type of pregonite with a somewhat extended and slender proboscis. I assume they are phylogenetically closer to the Egle longirostris species group than to E. concomitans .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Egle concomitans ( Pandellé, 1900 )
| Michelsen, Verner 2009 |
Egle concomitans (Pandellé)
| Hua, L. 2006: 98 |
| Ackland, D. M. & Merz, B. 2003: 210 |
| Griffiths, G. C. D. 2003: 2293 |
| Darvas, B. & Dely-Draskovits, A. & Ackland, D. M. & Jermy, T. 2001: 309 |
| Michelsen, V. & Bartak, M. 2001: 457 |
| Meixnerova, O. & Rozkosny, R. 1999: 383 |
| Teschner, D. 1999: 131 |
| Wei, L. & Xue, W. & Cui, C. 1998: 660 |
| Bartak, M. & Michelsen, V. & Rozkosny, R. 1990: 441 |
| Bartak, M. & Michelsen, V. & Rozkosny, R. 1989: 316 |
| Michelsen, V. 1988: 277 |
Lasiomma concomitans (Pandellé)
| Hennig, W. 1972: 429 |
Pegohylemyia concomitans (Pandellé)
| Hennig, W. 1967: 179 |
Chortophila (Nudaria) concomitans (Pandellé)
| Karl, O. 1928: 173 |
Hylemyia ithacensis
| Huckett, H. C. 1965: 66 |
| Huckett, H. C. 1924: 28 |
Chortophila cacuminata
| Hennig, W. 1972: 429 |
| Villeneuve, J. 1923: 95 |
Chortophila concomitans (Pandellé)
| Seguy, E. 1923: 123 |
Hylemyia hinei
| Malloch, J. R. 1920: 278 |
Anthomyia (Chortophila) concomitans Pandellé, 1900: 251
| Pandelle, L. 1900: 251 |