
Amphibioplana onnisi, Stocchino & Sluys, 2021
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlaa183 |
|
publication LSID |
lsid:zoobank.org:pub:35092390-CE13-4E2D-9C70-47014A4C165F |
|
DOI |
https://doi.org/10.5281/zenodo.5762057 |
|
persistent identifier |
https://treatment.plazi.org/id/D81287D8-0518-5E49-BDD5-A93EA48BE1A4 |
|
treatment provided by |
Plazi |
|
scientific name |
Amphibioplana onnisi |
| status |
sp. nov. |
AMPHIBIOPLANA ONNISI STOCCHINO & SLUYS , SP. NOV.
( FIGS 3–15 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 )
Z o o b a n k r e g i s t r a t i o n: l s i d: u r n:l s i d:z o o b a n k. org:act: 01B99CA0-9692-477F-8913-8A476638C57B
Diagnosis: As that of the genus.
Etymology: The specific epithet honours the Sardinian biospeleologist and co-author Carlo Onnis, who discovered and sampled all populations of the new species.
Material examined: Holotype: RMNH.VER. 19956.a, Grutta ‘e Pauli Cave (39°53’13.45”N, 9°20’39.47”E), terrestrial habitat, Seui, Sardinia, Italy, 18 January 2015, coll. C. Onnis, horizontal sections (hor. sect.) of anterior portion on one slide; sagittal sections (sag. sect.) of posterior portion on two slides.
Paratypes: CGAS Pla 19.1a, ibid., sag. sect. of anterior portion on two slides; CGAS Pla 19.1b, ibid., sag. sect. of posterior portion on two slides; CGAS Pla 19.2, ibid., sag. sect. on three slides; RMNH.VER. 19956.b, ibid., sag. sect. on two slides; RMNH.VER. 19956.c, ibid., sag. sect. on two slides; RMNH.VER. 19956.d, ibid., sag. sect. on two slides.
Other material: Grutta ‘e Pauli Cave (39°53’13.45”N, 9°20’39.47”E), Seui, Sardinia: CGAS Pla 20.1, freshwater habitat 11 October 2015, coll. C. Onnis, sag. sect. on two slides; CGAS Pla 20.2, ibid., sag. sect. on two slides; CGAS Pla 20.3, ibid., sag. sect. on three slides; CGAS Pla 20.4, ibid., sag. sect. on three slides; CGAS Pla 20.5, ibid., sag. sect. on two slides; RMNH. VER. 19957.a, ibid., sag. sect. on two slides; CGAS Pla 20.6, ibid., sag. sect. on two slides; RMNH.VER. 19957.b, ibid., sag. sect. on two slides, RMNH.VER. 19957.c, ibid., sag. sect. on two slides, RMNH.VER. 19957.d, ibid., sag. sect. of anterior portion on two slides; RMNH.VER. 19957.e, ibid., hor. sect. of posterior portion on one slide. CGAS Pla 21, freshwater habitat, 26 March 2018, coll. C. Onnis, six entire specimens fixed in Bouin’s fluid and preserved in ethanol.
Istirzili Cave (40°04’48.29”N, 9°37’12.50”E), Baunei, Sardinia: RMNH.VER. 19958.a, freshwater habitat, 22 January 2017, coll. C. Onnis, sag. sect. on two slides; CGAS Pla 22.1, ibid., sag. sect. on three slides; CGAS Pla 22.2, ibid., sag. sect. on two slides; CGAS Pla 22.3, ibid., sag. sect. on two slides; RMNH.VER. 19958.b, ibid., sag. sect. on one slide; CGAS Pla 22.4, ibid., sag. sect. on one slide; RMNH.VER. 19958.c, ibid., sag. sect. on three slides; RMNH.VER. 19958.d, ibid., sag. sect. on one slide; RMNH.VER. 19958.e, ibid., sag. sect. on two slides; RMNH.VER. 19958.f, ibid., hor. sect. on one slide; CGAS Pla 22.5, ibid., hor. sect. on two slides; CGAS Pla 22.6, ibid., transverse sections (transv. sect.) on five slides.
San Pietro Cave (39°24’27.38”N, 8°29’35.40”E), Fluminimaggiore, Sardinia: CGAS Pla 23, terrestrial habitat, 26 June 2015, coll. C. Onnis, sag. sect. on one slide. RMNH.VER. 19959.a, freshwater habitat, 5 October 2015, coll. C. Onnis, sag. sect. on one slide. CGAS Pla 24.1, freshwater habitat, May 2016, coll. C. Onnis, sag. sect. on one slide; RMNH.VER. 19960.a, ibid., sag. sect. on one slide; CGAS Pla 24.2, ibid., sag. sect. on one slide; RMNH.VER. 19960.b, ibid., sag. sect. on one slide; CGAS Pla 24.3, ibid., sag. sect. on one slide; RMNH. VER. 19960.c, ibid., sag. sect. on one slide; CGAS Pla 24.4, ibid., sag. sect. on one slide; CGAS Pla 24.5, ibid., sag. sect. on one slide; CGAS Pla 24.6, ibid., sag. sect. on one slide; CGAS Pla 24.7, ibid., sag. sect. on one slide; RMNH.VER. 19960.d, ibid., sag. sect. on one slide; CGAS Pla 24.8, ibid., sag. sect. on one slide; CGAS Pla 24.9, ibid., sag. sect. on one slide; CGAS Pla 24.10, ibid., sag. sect. on two slides; CGAS Pla 24.11, ibid., sag. sect. on two slides; CGAS Pla 24.12, ibid., sag. sect. on two slides; CGAS Pla 24.13, ibid., sag. sect. on two slides; CGAS Pla 24.14, ibid., sag. sect. on two slides; CGAS Pla 24.15, ibid., sag. sect. on two slides; RMNH.VER. 19960.e, ibid., sag. sect. on one slide; CGAS Pla 24.16, ibid., sag. sect. on one slide; CGAS Pla 24.17, ibid., sag. sect. on one slide; CGAS Pla 24.18, ibid., sag. sect. on one slide; CGAS Pla 24.19, ibid., sag. sect. on one slide; CGAS Pla 24.20, ibid., sag. sect. on one slide; RMNH.VER. 19960.f, ibid., sag. sect. on one slide; CGAS Pla 24.21, ibid., sag. sect. on two slides; CGAS Pla 24.22, ibid., sag. sect. on one slide; RMNH.VER. 19960.g, ibid., hor. sect. on one slide; CGAS Pla 24.23, ibid., hor. sect. on one slide; RMNH.VER. 19960.h, ibid., transv. sect. on four slides.
Description
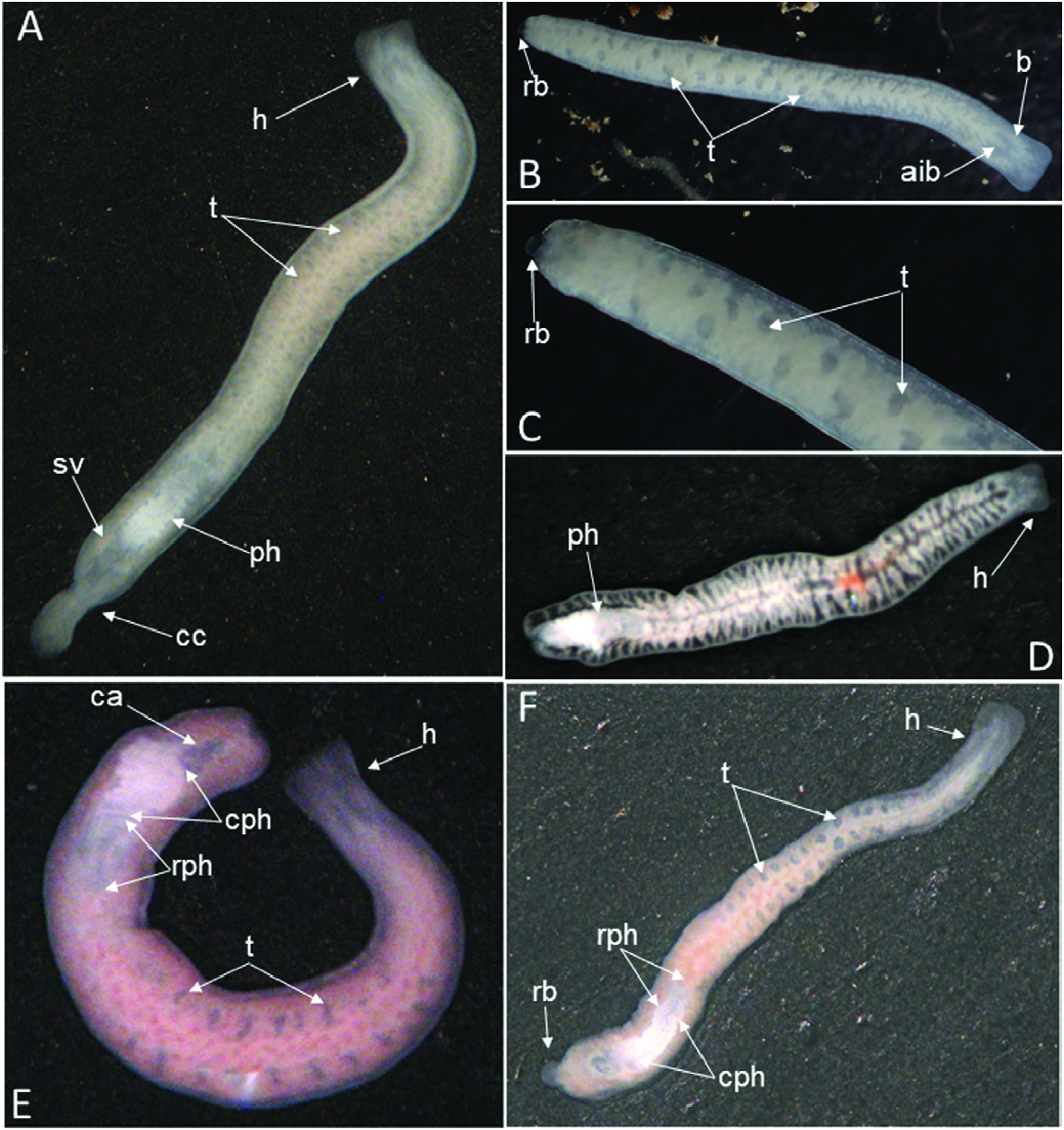
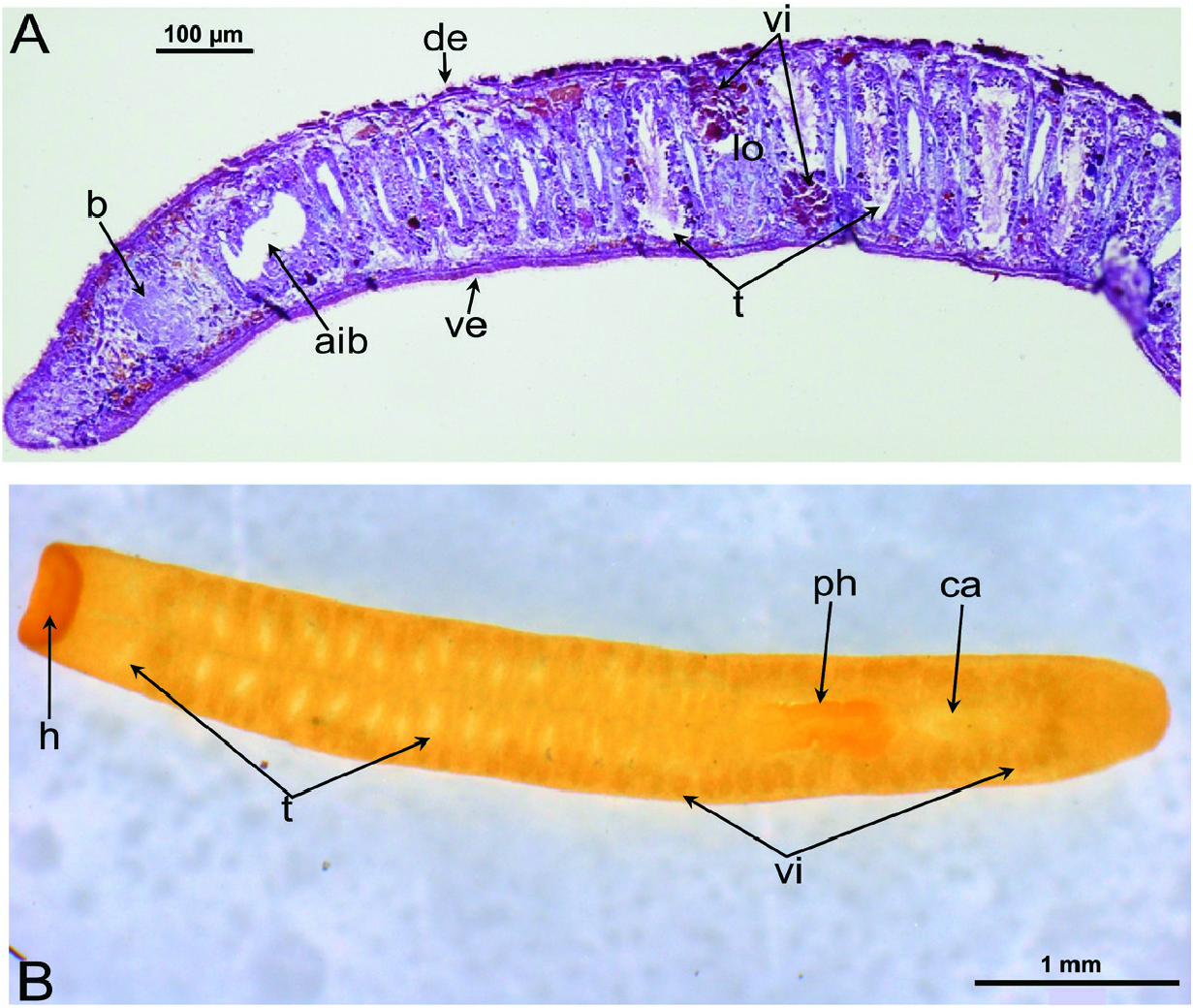
External features: Body size of live specimens ranges from 4 to 6 mm long and 0.4–0.8 mm wide. Preserved specimens measured up to 3.5–4.0 mm long and 0.4– 0.8 mm wide. The animals are unpigmented and fairly transparent in vivo, so that the intestinal branches, brain, testes, pharynx and copulatory apparatus are clearly visible. The truncated head is devoid of eyes; posterior end rounded ( Fig. 5 View Figure 5 ).
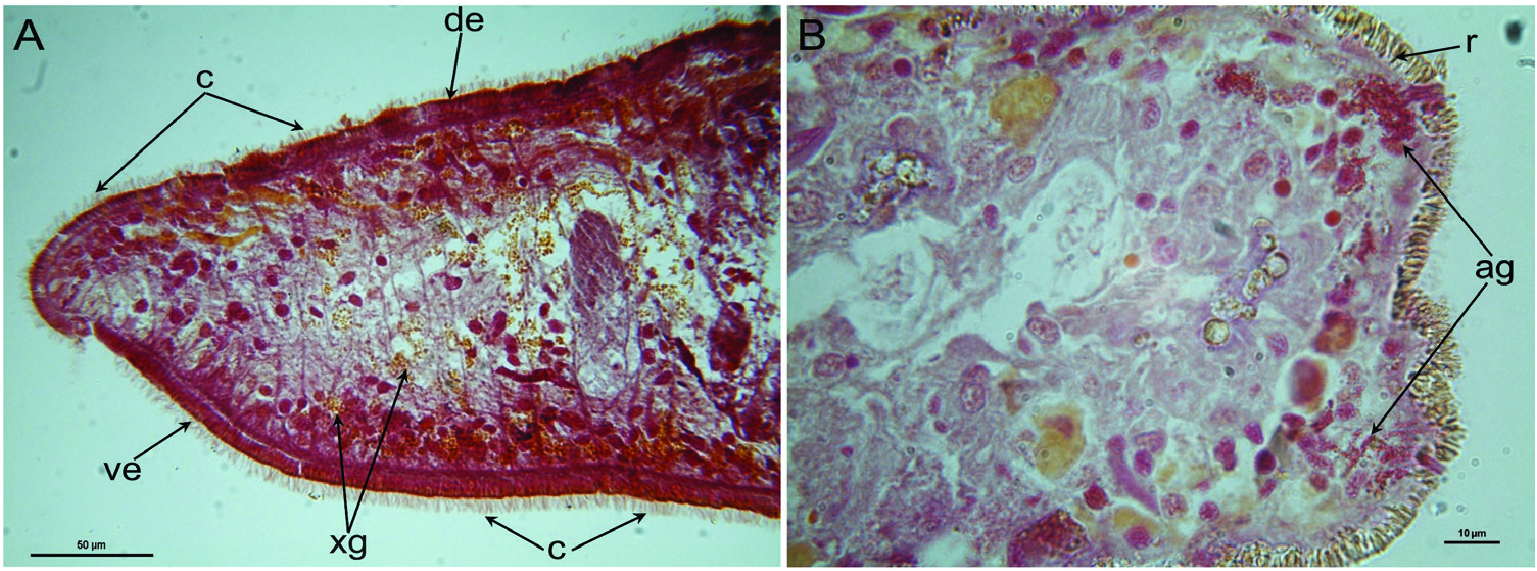
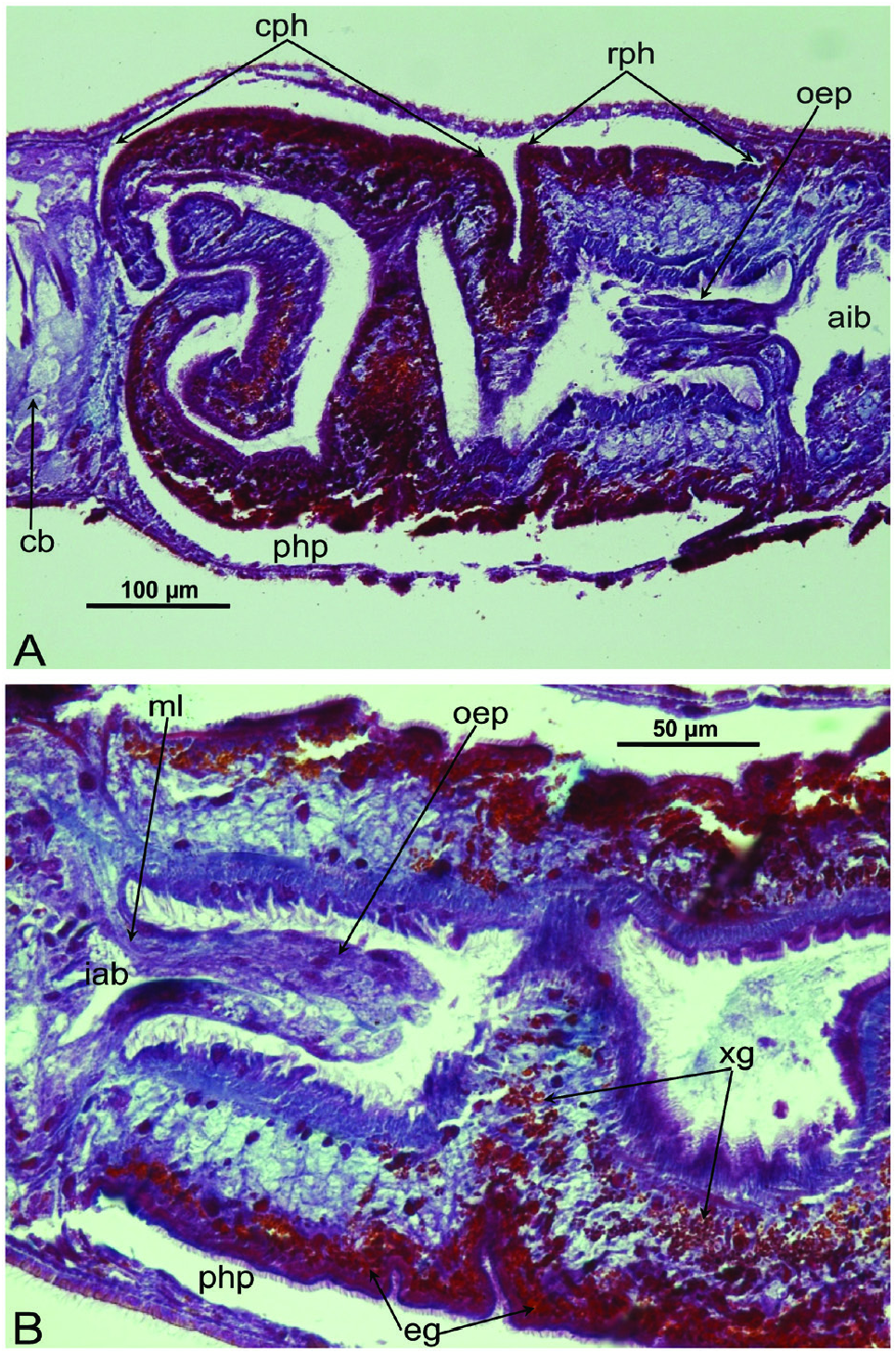
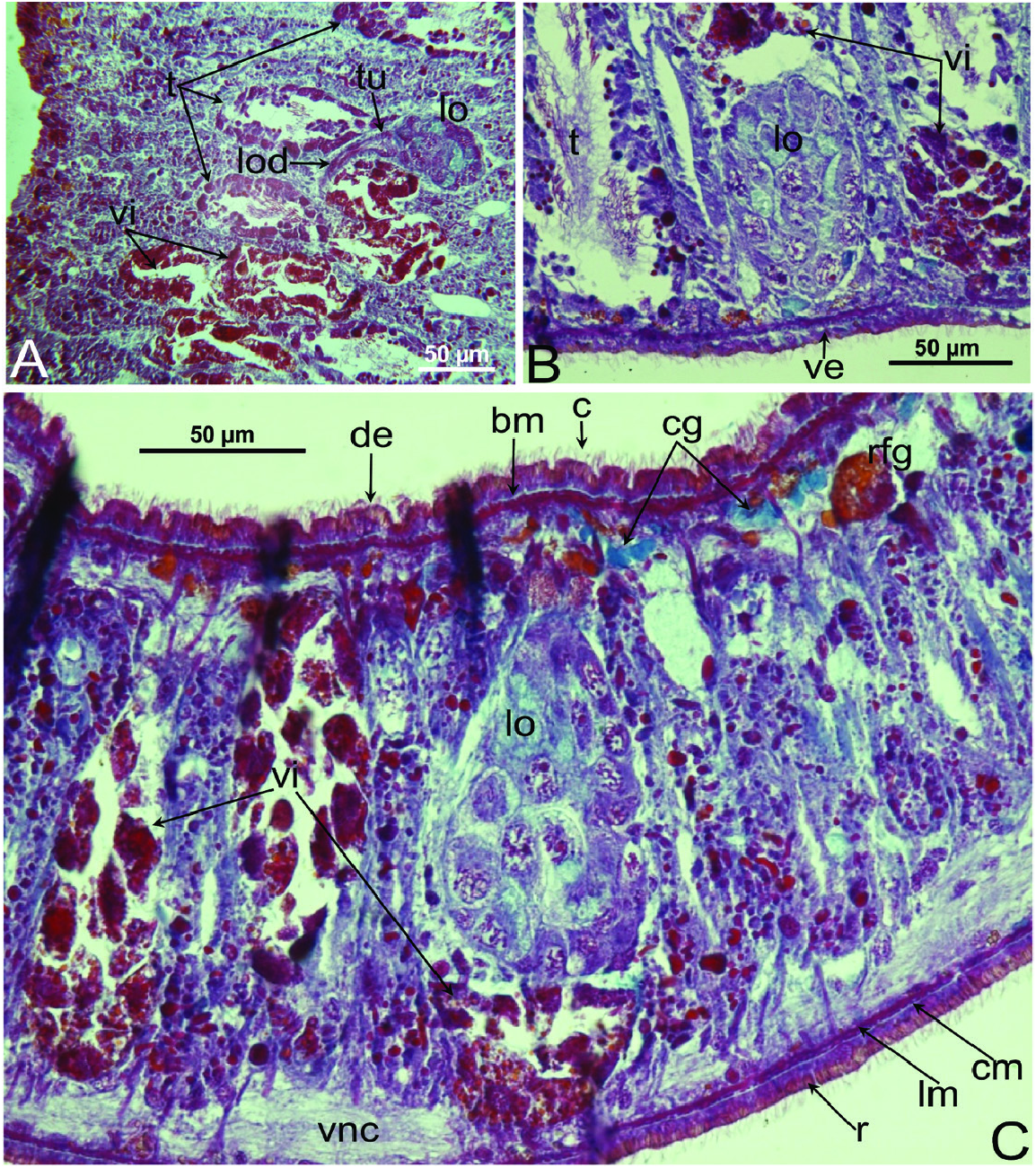
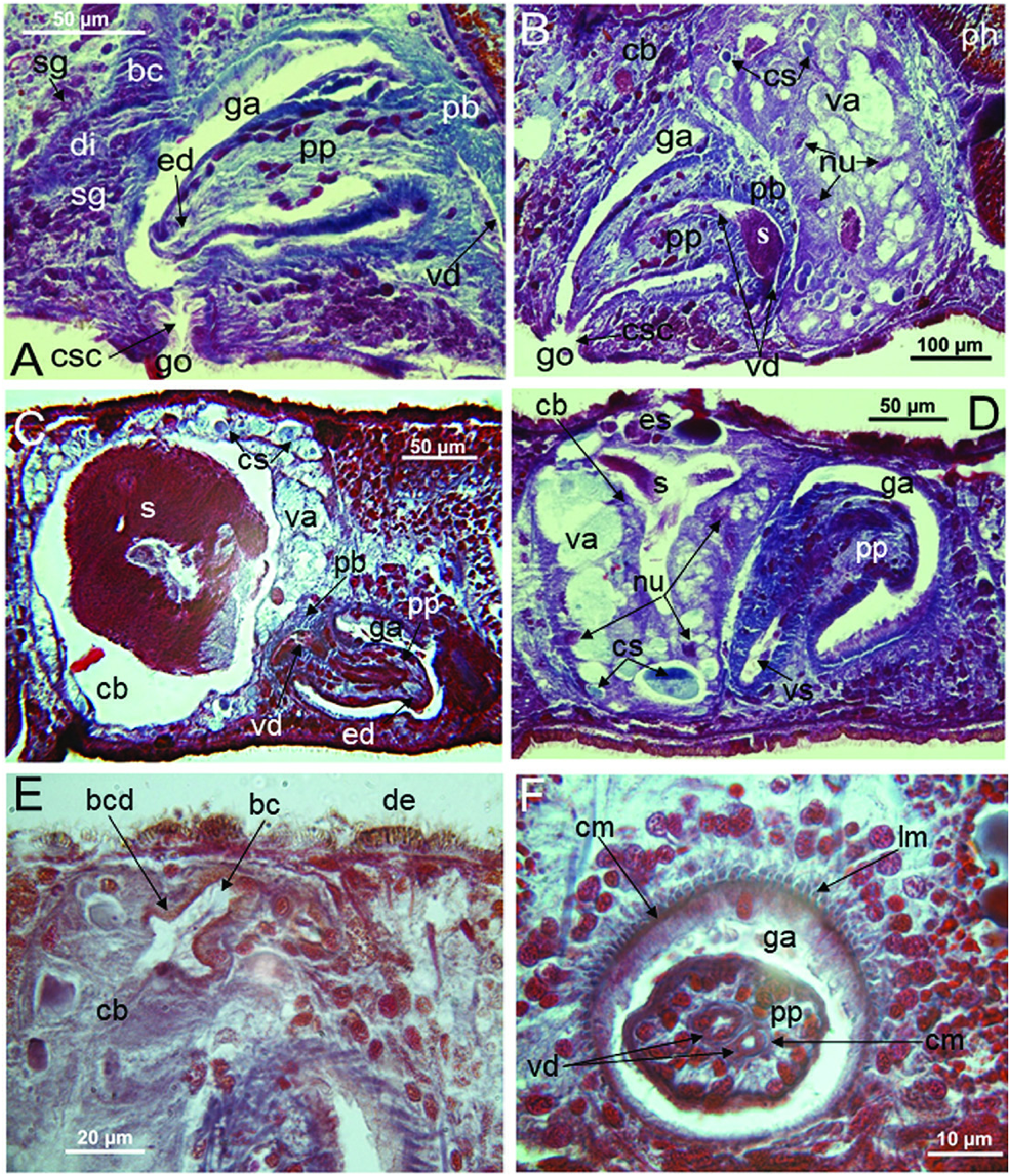
Epidermis and cutaneous musculature: The entire epidermis is constituted of a cuboidal, nucleated epithelium that is completely clothed in cilia that have the same length on both dorsal and ventral body surface; dorsal and ventral epidermis have the same height. Rhabdites are abundant and uniformly distributed on the entire body surface, with erythrophil, rhabditeforming glands being located in the parenchyma underneath the subepidermal musculature ( Figs 6A View Figure 6 , 7B View Figure 7 , 10C View Figure 10 ). Dorsal and ventral rhabdites have the same height. The epidermis also receives a coarse granular, xanthophil secretion from glands that are mainly located in the anterior region of the body, beneath the subepidermal musculature, with several glands also lying scattered in the parenchyma ( Figs 6A View Figure 6 , 7A View Figure 7 ). The epidermis receives also an amorphous secretion from cyanophil glands located beneath the subepidermal musculature ( Fig. 10B, C View Figure 10 ). Underneath the epidermis, the basement membrane is evident as a pale blue, amorphous line ( Fig. 10C View Figure 10 ). The openings of adhesive glands, producing a fine, granular, erythrophil secretion, are concentrated at the front end and at the posterior tip of the body ( Figs 6B View Figure 6 , 7A View Figure 7 ). Subepidermal musculature consists of only two thin layers: an outer, subepidermal layer of circular muscles and an inner layer of longitudinal muscles ( Figs 10B, C View Figure 10 , 11 View Figure 11 , 12 View Figure 12 ). Dorsal and ventral musculatures show the same thickness.
Nervous system: In horizontal sections, the bilaterally symmetrical brain has the shape of an ‘H’ and lies on top of the two ventral nerve cords, which run in the caudal direction along the length of the animal ( Figs 7A View Figure 7 , 10C View Figure 10 ). The central neuropil of the brain is surrounded by an outer layer of neuronal cell bodies ( Fig. 7A View Figure 7 ). In none of the specimens examined was it possible to discern structures related to an auricular sense organ, in view of the fact that the epidermal cells are of a uniform height and are all provided with both cilia and rhabdites.
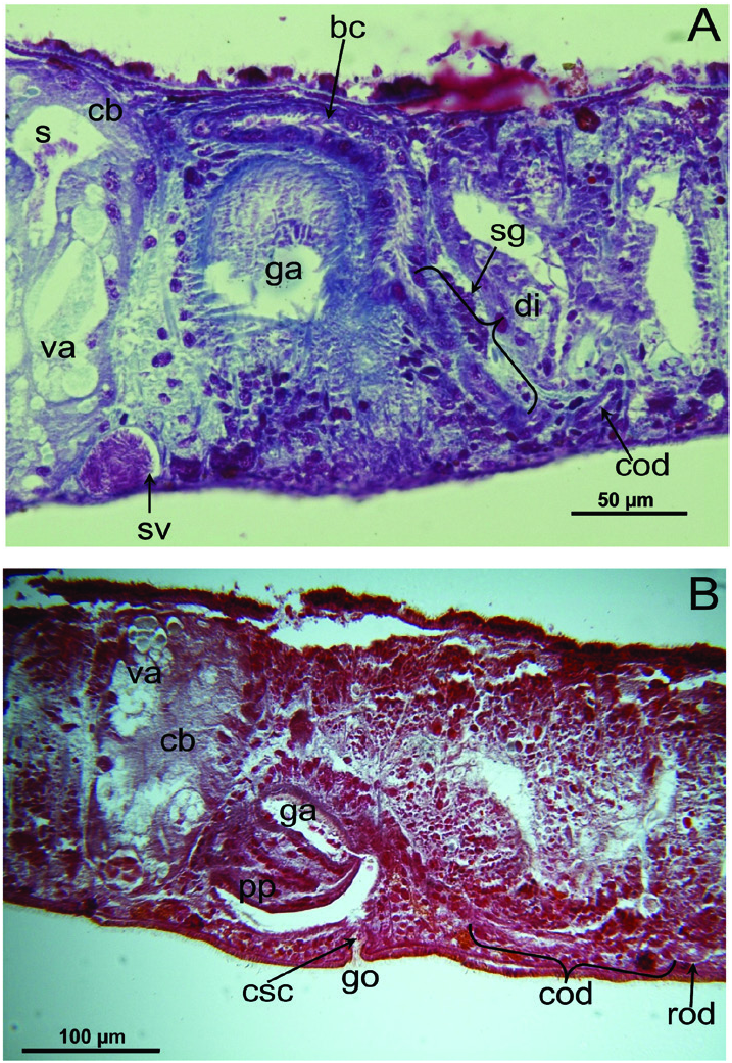
Pharynx and intestine: The unpigmented, plicate pharynx is located in the posterior half of the body and measures ~one-tenth of the body length, at least when it is situated within the pharyngeal pocket. In point of fact, the actual size of the pharynx must be greater than this as it is considerably folded and convoluted at its median and distal tracts ( Figs 5F View Figure 5 , 8 View Figure 8 ). In specimens from Grutta ‘e Pauli Cave and Istirzili Cave the proximal tract of the pharynx, corresponding to one-third of its total length, has a rectilinear course and at its root it is provided with a kind of oesophagal protrusion that actually projects into the pharynx lumen, instead of the gut. In specimens from San Pietro Cave the pharynx has a less convoluted shape, while the oesophagal protrusion is smaller than in specimens of the other two populations. This oesophagal protrusion is constituted by gut tissue traversed for almost its entire length by thin, longitudinal muscle fibres ( Fig. 8B View Figure 8 ). The epithelium lining the proximal, anterior part of the pharynx lumen is provided with long cilia. The muscle coat underneath the outer pharynx epithelium is bilayered, i.e. constituted by a subepithelial layer of longitudinal muscle, followed by a layer of circular muscles. The musculature underneath the epithelium lining the pharynx lumen is composed of a thick, subepithelial layer of circular muscles, followed by a layer of longitudinal muscles. Two types of pharyngeal glands are evident: one type producing a fine-grained erythrophil secretion and the other a coarse-grained xanthophil secretion. In the majority of specimens, the mouth opening is located at the posterior end of the pharyngeal pouch, but it may be shifted anteriad, so that in some cases it is located at ~one-sixth, and in one case at ~one-third, of the distance between the posterior end of the pharyngeal pouch and the root of the pharynx. The intestine is of the usual tripartite shape, with the anterior intestinal trunk terminating just behind the brain. Posterior intestinal branches unite behind the copulatory apparatus to form a single branch ( Figs 5 View Figure 5 , 7 View Figure 7 , 9A View Figure 9 ).
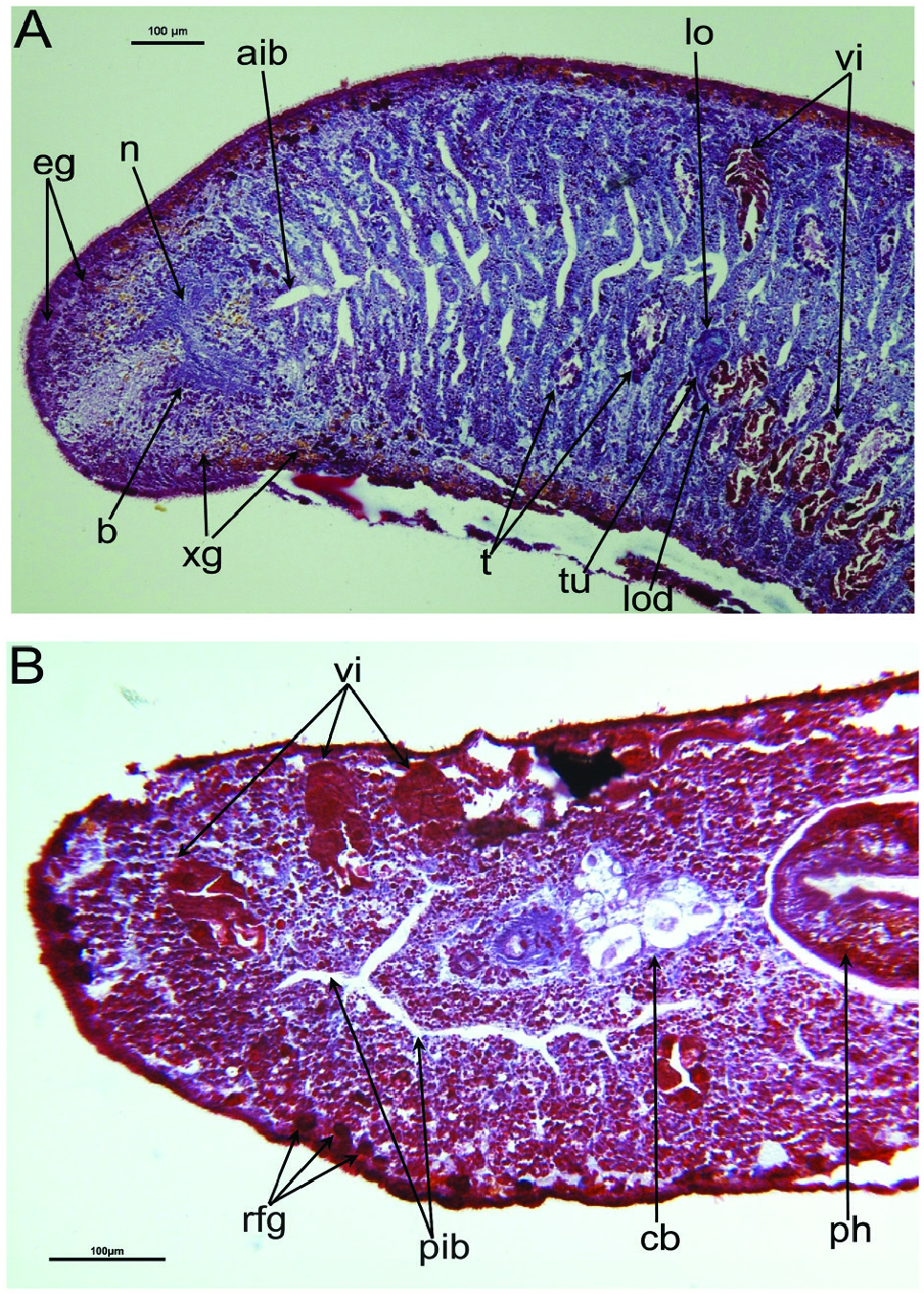
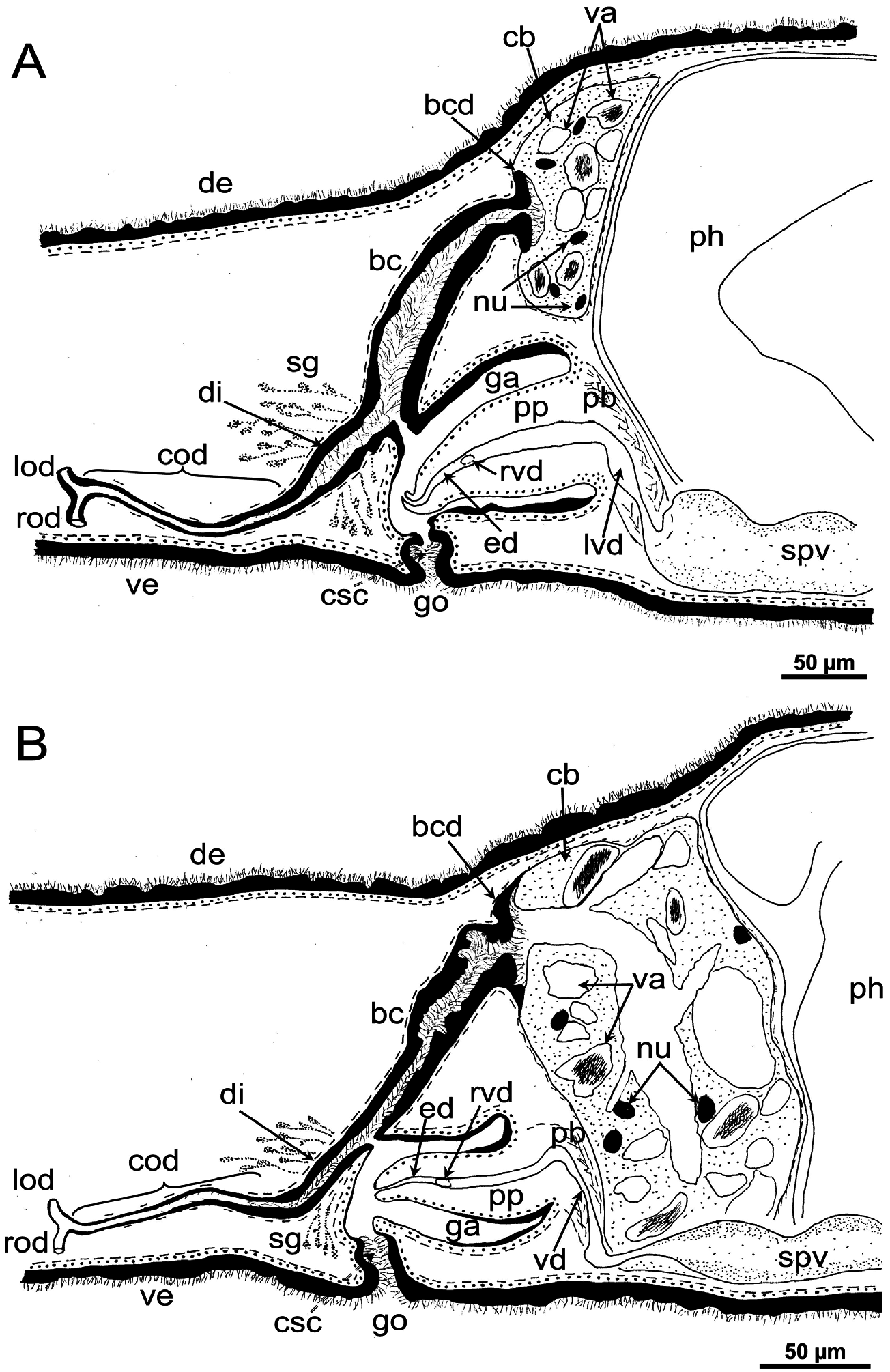
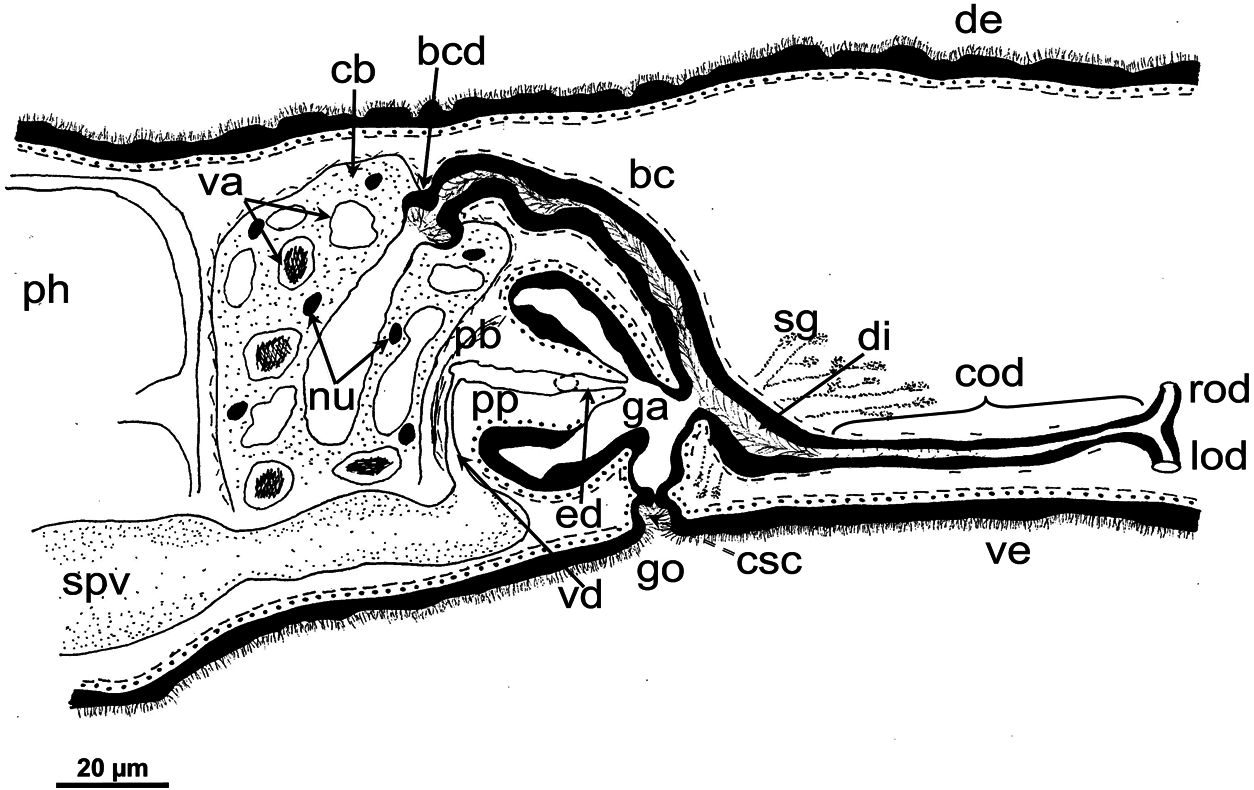
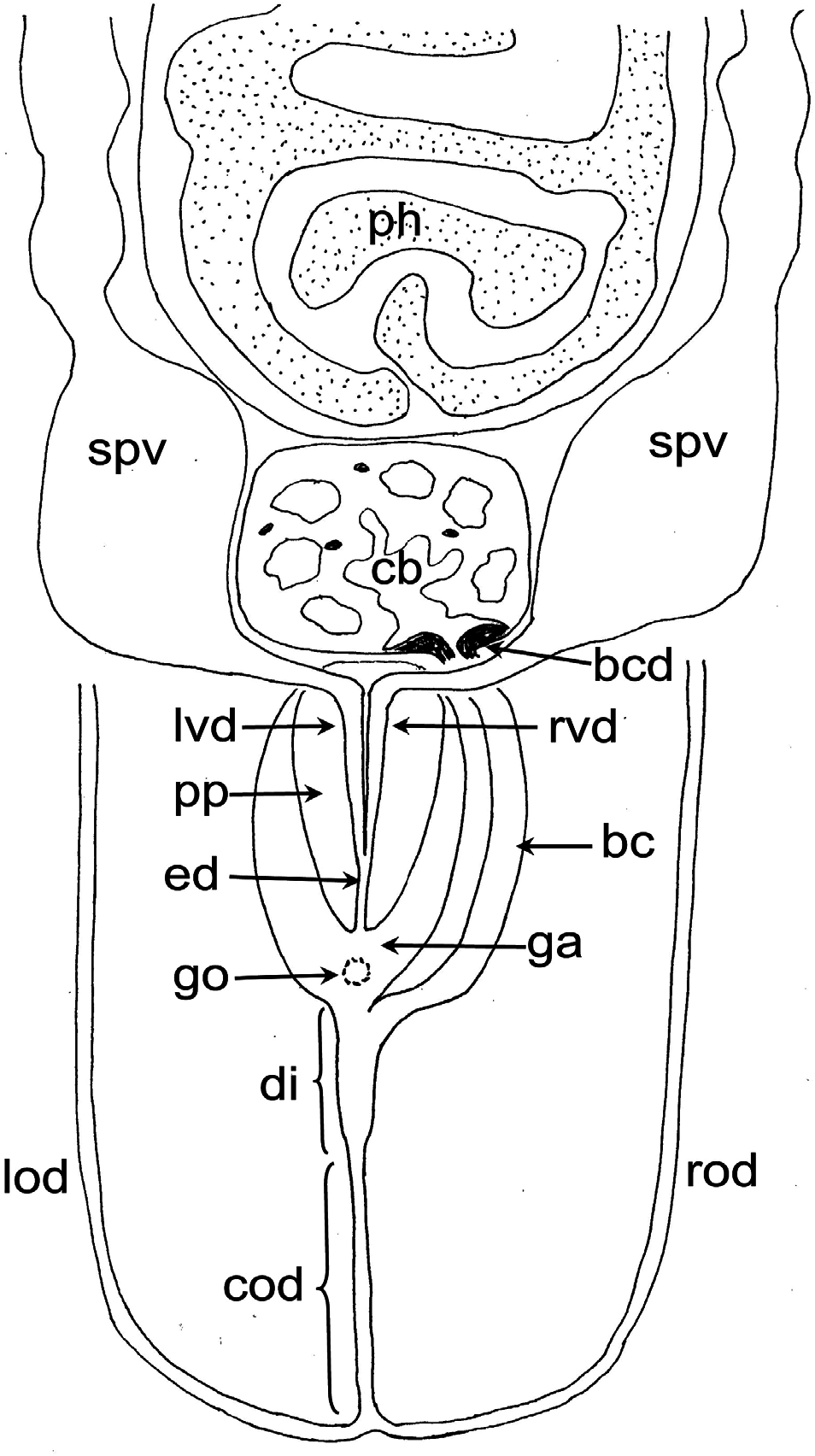
Female reproductive system: The two ovoid ovaries are located ventrally, far behind the brain and just behind the most anterior pair of testes. The female gonads are located at ~one-third to one-quarter of the distance between the brain and the root of the pharynx, i.e. at ~one-eighth of the distance between the brain and the posterior end of the body. The ovaries occupy one-half (specimen RMNH.VER. 19956.d) or three-quarters (specimen CGAS Pla 19.2) of the dorsoventral space. The anterior portion of each oviduct is expanded to form a tuba, which communicates with the dorsolateral part of the corresponding ovary ( Figs 9A View Figure 9 , 10 View Figure 10 ). The oviducts run ventrolaterally to the vitellaria and laterally to the ventral nerve cords, and extend posteriorly to beyond the level of the genital pore and up to the level of the penultimate vitellarium. Hereafter, the oviducts bend medially and subsequently unite to form a long common oviduct. The common oviduct runs anteriad and then communicates with the posterior diverticulum of the bursal canal ( Figs 7A View Figure 7 , 9A View Figure 9 , 10–13 View Figure 10 View Figure 11 View Figure 12 View Figure 13 , 15 View Figure 15 ). Oviducts and common oviduct are lined with an epithelium consisting of cuboidal, nucleated and ciliated cells. The posterior part of the common oviduct is surrounded by a thin layer of longitudinal muscles that becomes even less distinct towards the point of fusion of the two oviducts. We were unable to discern muscles around the oviducts so that, apparently, these ducts are devoid of any musculature ( Figs 11 View Figure 11 , 12 View Figure 12 , 15 View Figure 15 ). The lumen of the common oviduct, as well as that of the oviducts, is always well discernible. Vitellaria are situated laterally to the testes and between the intestinal branches, extending from the level of the ovaries to behind the copulatory apparatus; they are well developed and occupy almost the entire dorsoventral space of the body ( Figs 7 View Figure 7 , 9 View Figure 9 , 10 View Figure 10 ). The copulatory bursa, located just behind the pharyngeal pouch, is lined by a glandular and highly vacuolated, syncytial epithelium with scattered nuclei. Many of the vacuoles are richly provided with an amorphous, erythrophil and/or cyanophil secretion and in some cases contain sperm ( Figs 7B View Figure 7 , 11–13 View Figure 11 View Figure 12 View Figure 13 , 14B–E View Figure 14 , 15 View Figure 15 ). In the majority of the specimens, the bursal lumen contains a mass of free sperm and, in some cases, is so voluminous that it noticeably compresses the lining epithelium of the bursa (e.g. specimen RMNH.VER. 19958.a; see Fig. 14C View Figure 14 ). The bursa is surrounded by a layer of muscles. The size of the copulatory bursa varies between specimens, but in the majority of cases it is large and occupies the entire dorsoventral space, while occasionally it occupies only the dorsal half of the body space. The bursal canal originates from the posterior wall of the genital atrium and immediately thereafter gives rise to a posterior expansion that forms a kind of diverticulum, which receives the secretion of erythrophil shell glands and communicates with the common oviduct. From thereon, the bursal canal curves anterodorsad, while running, usually, to the right side of the rest of the copulatory apparatus and then communicates with the copulatory bursa. For a short distance, the bursal canal protrudes into the bursa, thus forming a kind of valve or diaphragm ( Figs 11– 13 View Figure 11 View Figure 12 View Figure 13 , 14E View Figure 14 ). The bursal canal is lined with an epithelium consisting of cuboidal or cylindrical, nucleated cells and is surrounded by a layer of longitudinal muscles ( Figs 11 View Figure 11 , 12 View Figure 12 , 15A View Figure 15 ). The epithelial cells of the bursal canal, which are provided with long cilia, produce an erythrophil secretion that is discharged into the lumen. The point of transition between bursal canal diverticulum and common oviduct is not always clear-cut, but can be recognized by the fact that the common oviduct has a smaller diameter and is lined with a lower epithelium, provided with shorter cilia, and is surrounded by a thinner coat of muscles. In addition, oviducts and common oviduct stain less strongly than the bursal canal diverticulum.
Male reproductive system: The well-developed testes are arranged in two regular rows, one row on either side of the body, with the follicles occupying the entire dorsoventral space. The most anterior testes are situated far behind the brain, albeit anteriorly to the ovaries, and from thereon extend caudally, with the most posterior follicles located far (~ 500–550 µm) from the root of the pharynx ( Figs 5 View Figure 5 , 6A View Figure 6 , 9 View Figure 9 , 10 View Figure 10 ). The vasa deferentia run dorsally to the ventral nerve cords and already in the prepharyngeal part of the body they form voluminous spermiducal vesicles that are packed with sperm, at least in sexually mature specimens ( Figs 5A View Figure 5 , 11–13 View Figure 11 View Figure 12 View Figure 13 , 15A View Figure 15 ). The small and sometimes indistinct penis bulb is generally located just behind the copulatory bursa. However, in those specimens in which the bursa is small, the penis bulb almost touches the caudal wall of the pharyngeal pouch. At the level of the penis bulb, the spermiducal vesicles notably decrease in diameter and hereafter they curve dorsad and then abruptly penetrate the anteroventral wall of the penis bulb, after which they symmetrically ascend to reach the level of the medial part of the penis. After a while, the ducts curve again abruptly or via a knee-shaped bend and then turn posteriad, after which they run parallel and close to each other through the central portion of the penis papilla. At ~three-quarters of the length of the papilla, the vasa deferentia fuse to form a short ejaculatory duct, which runs a central course through the remainder of the penis papilla and opens at its tip ( Figs 11–13 View Figure 11 View Figure 12 View Figure 13 , 14A–D, F View Figure 14 , 15B View Figure 15 ). The vasa deferentia are lined with an epithelium consisting of low, cuboidal and nucleated cells and are surrounded by a layer of circular muscle. The ejaculatory duct is lined with cuboidal, nucleated cells. The symmetrical, conical penis papilla is covered by a flat infranucleated epithelium, which is underlain by a subepithelial layer of circular muscle ( Figs 11–14A–D, F View Figure 11 View Figure 12 View Figure 13 View Figure 14 , 15B View Figure 15 ). Weak, finely granular, erythrophil secretions were discerned in the penis papilla. The genital atrium, which houses the penis papilla, is lined by an epithelium consisting of, cuboidal-columnar, infranucleated cells with a weak erythrophil secretion that is underlain by a subepithelial layer of circular muscle, followed by a layer of longitudinal muscle fibres ( Figs 11–14A–D, F View Figure 11 View Figure 12 View Figure 13 View Figure 14 , 15 View Figure 15 ). In all specimens examined, the ventral gonopore directly leads to a cup-shaped chamber, which is formed by an invagination of the ventral epidermis, as it is lined by a ciliated epithelium that is provided with rhabdites and a subepidermal layer of circular muscles. The dorsal wall of this chamber is provided with a small dorsal pore through which it communicates with the atrium ( Figs 11 View Figure 11 , 12 View Figure 12 , 14A, B View Figure 14 , 15B View Figure 15 ). Cement glands opening into the gonopore or the chamber were not discerned.
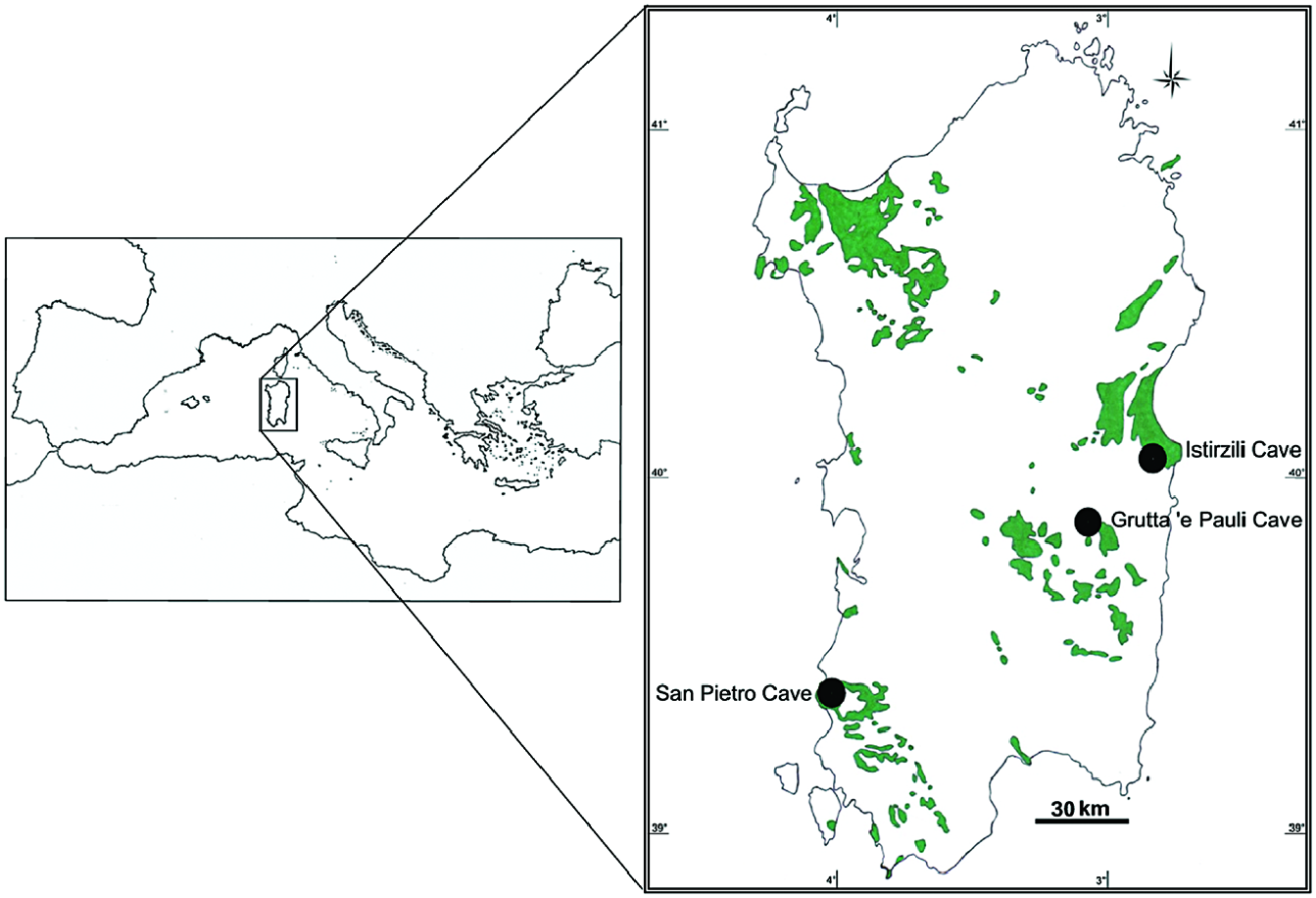
Geographical distribution: Exclusively known from three karstic caves in Sardinia, Italy ( Fig. 1 View Figure 1 ).
Habitat: Planarians were found in three karstic caves, characterized by intermittent presence of dripping water. These caves are located in three karstic massifs that are greatly isolated from each other, representing distinct hydrological systems ( Cassola, 1982).
The Grutta ‘e Pauli Cave is located at an elevation of ~ 1025 m above sea level (a.s.l.) in the karstic Mesozoic (Jurassic) massif of the Tacchi d’Ogliastra in the central-eastern part of Sardinia ( Figs 1 View Figure 1 , 4A View Figure 4 ). Specimens were first collected at ~ 40 m from the entrance from under stones that were firmly embedded in humid, clayey soil. Some individuals were found adhering to the stones, while others were found on the clayey soil, thus in a basically terrestrial habitat. Animals were motionless and tightly curled up ( Fig. 3 View Figure 3 ). However, after only having been touched with a small paint brush they started to move, after which the worms were transferred to a vial with water in which, subsequently, they behaved as freshwater planarians, exhibiting the gliding manner of locomotion. Subsequent to this first collection, animals were found also in temporary pools, formed by the dripping water and provided with a clayey bottom, at ~ 50 m from the entrance of this cave ( Fig. 4B View Figure 4 ); associated fauna was exclusively Ostracoda.
The San Pietro Cave is located at an elevation of ~ 225 m a.s.l. at the south-western coast of Sardinia, in the Iglesiente region, which is considered to be the most ancient karst of Italy, dating back to the Cambrian ( Fig. 1 View Figure 1 ). Flatworms were found under stones that were firmly embedded in a humid, clayey substrate, at ~ 50 m from the entrance, as well as in some rimstone basins or gours, with a muddy bottom, at ~ 260 m from the entrance of the cave ( Fig. 4C View Figure 4 ). While under the stones, the worms were motionless and tightly curled up.
The Istirzili Cave is located at an elevation of ~ 507 m a.s.l. in the karstic Mesozoic (Jurassic) highland massif of the Supramonte of Baunei in the centraleast of Sardinia ( Fig. 1 View Figure 1 ). Planarians were found only in a small temporary pool located near (~ 5 m) the cave entrance, with a clay bottom characterized by the presence of small stones and leaves ( Fig. 4D View Figure 4 ).
Reproduction: The great majority of individuals were sexual at collection, but cocoons were not found in any of the three caves. Furthermore, signs of presumably asexual reproduction by fission were displayed by some sexually mature specimens from the population from Grutta ‘e Pauli Cave. These animals showed five different conditions: (1) a transverse constriction (some individuals) just caudally to the copulatory apparatus, suggesting a preliminary phase of fission ( Fig. 5A View Figure 5 ); (2) a small, caudal regenerating blastema (one individual), suggesting a previous process of prepharyngeal fission ( Fig. 5B, C View Figure 5 ); (3) a body in which the caudal tip was located just behind the pharynx, suggesting a recent postpharyngeal fission (one individual) ( Fig. 5D View Figure 5 ); (4) a body in which the caudal end was located just behind the copulatory apparatus, suggesting a presumably recent fission (one individual) just caudally to the copulatory apparatus ( Fig. 5E View Figure 5 ); (5) a small, regenerating blastema at the current tail end, suggesting a previous fission (one individual) just caudally to the copulatory apparatus ( Fig. 5F View Figure 5 ).
Movement and locomotion: Similar to typical freshwater planarians, the newly collected Sardinian animals characteristically exhibited a gliding motion over the substrate. However, during locomotion the worms tend to adhere temporarily to the substrate by their posterior end. The phase of adhesion was characterized by the anterior part of the animal moving rapidly with sideways swinging movements, as if exploring the surrounding environment, after which the animal releases itself from the substrate and resumes its gliding motion.


| C |
University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.