
Masirana suzukii, Ballarin & Eguchi, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5213.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:B5C1E409-DF40-4BDE-9702-DE79C3A252A9 |
|
DOI |
https://doi.org/10.5281/zenodo.7383527 |
|
persistent identifier |
https://treatment.plazi.org/id/D92987FD-6767-FFC9-BCC7-FD3CFF07F816 |
|
treatment provided by |
Plazi |
|
scientific name |
Masirana suzukii |
| status |
sp. nov. |
Masirana suzukii sp. nov.
(Japanese name: tiragamahinamashiragumoティラガマヒナマシラグモ)
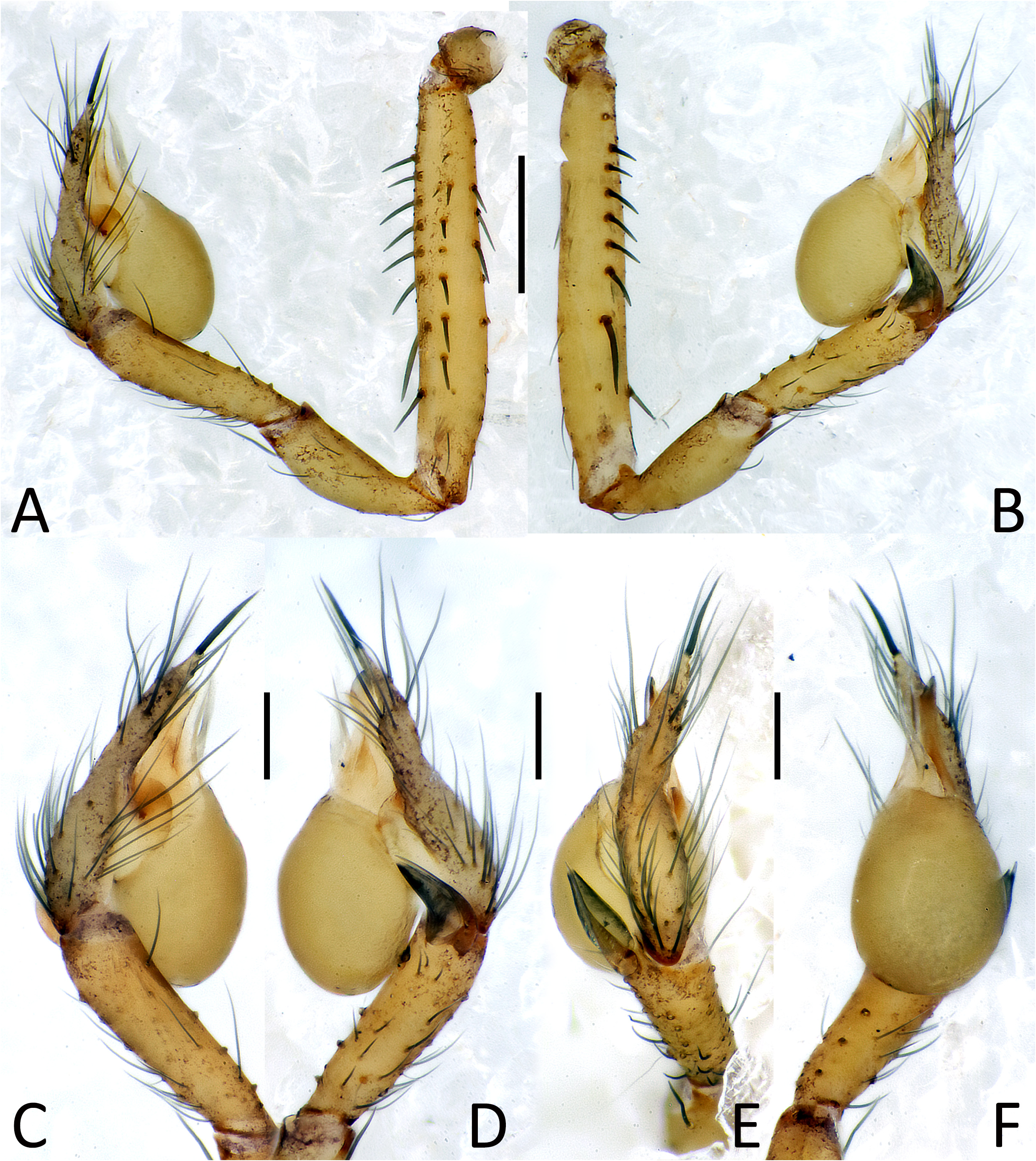
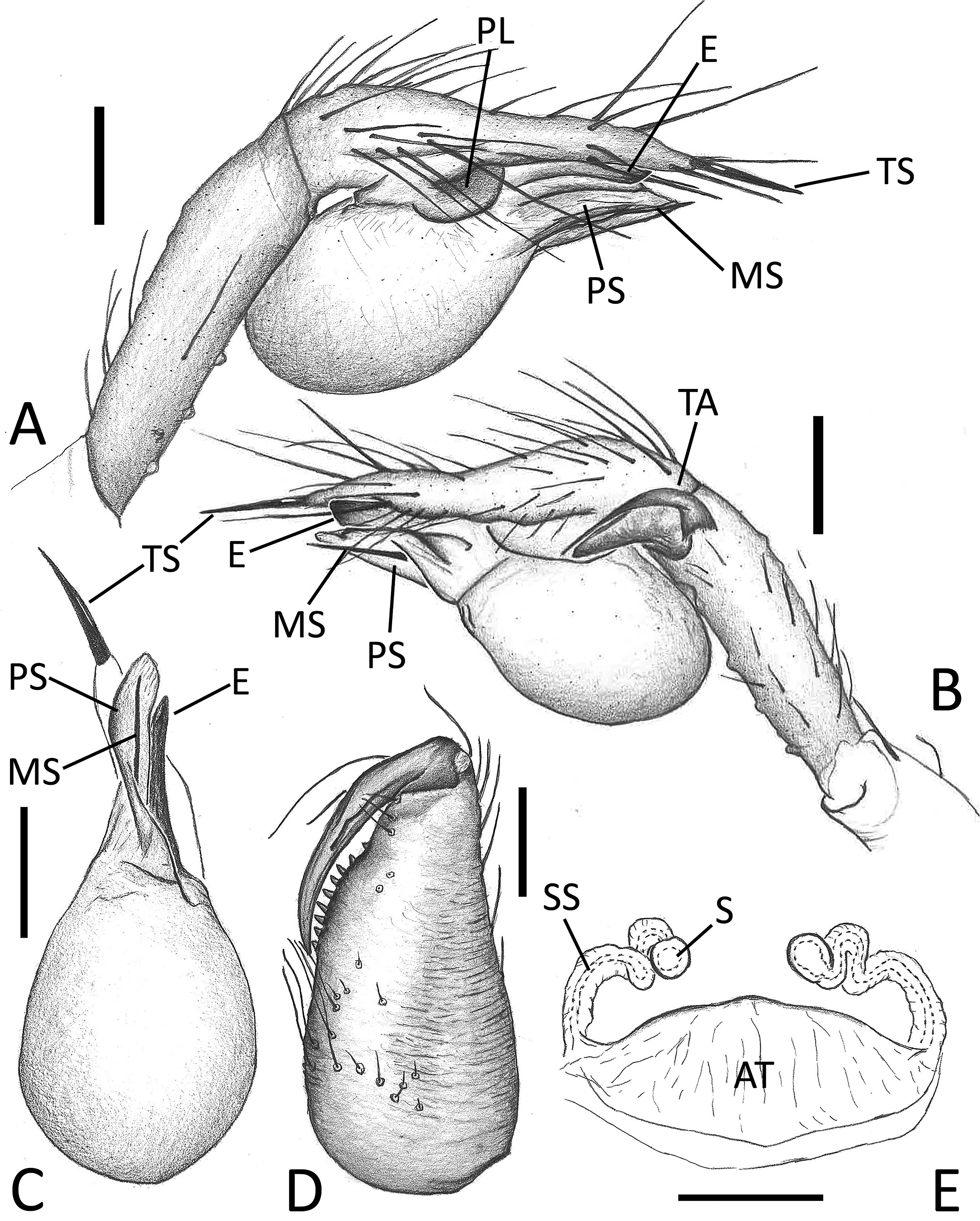
Figs. 4A–F View FIGURE 4 , 5A–E View FIGURE 5 , 6A–E View FIGURE 6 .
Material examined. ♂ Holotype. JAPAN: Okinawa Pref.: Okinawa-honto Is., Kunigami-gun, Nakijin-son, Tiragama cave (ティラガマ = Tametomo no horaana, Ẫ朝の洞Ẉ), 26.684°N, 128.005°E, 55 m a.s.l., short and rather humid cave, 17 November 2020, F. Ballarin leg. (NSMT-Ar 22246). GoogleMaps
Paratypes. JAPAN - same data as the holotype, 2♂, 1♀ GoogleMaps (2♂, 1♀ RMUF; 1♂, 1♀ FBPC) .
Etymology. The new species is a patronym in honor to our colleague and friend Yuya Suzuki (United Graduate School of Agricultural Sciences, Kagoshima University, Japan). Yuya is a young and promising arachnologist actively working on ethology, ecology and taxonomy of Japanese spiders, including cave species from the Ryukyus.
Diagnosis. Species closely related to Masirana changlini (Zhu & Tso, 2002) from Taiwan. The male of M. suzukii sp. nov. can be distinguished from the male of M. changlini by the different number of denticles on the chelicera (a single row of 9 denticles on the promargin in M. suzukii sp. nov. vs. two rows of denticles, 6 on the promargin and 7 on the retromargin, in M. changlini ; cf. Fig. 6D View FIGURE 6 vs. fig. 3 in Zhu & Tso 2002) and by the slimmer and longer tip of the cymbium bearing a short, stronger spine (vs. a shorter, tougher tip of cymbium lacking any thick apical spine in M. changlini ; cf. Figs. 4B, C–F View FIGURE 4 , 6A, B View FIGURE 6 vs. figs. 4, 5 in Zhu & Tso, 2002). In addition, the embolus of the new species shows a more rounded apex in contrast with a pointy apex in M. changlini (cf. Figs. 4B View FIGURE 4 , 6B View FIGURE 6 vs. fig. 4 in Zhu & Tso 2002). The male of M. suzukii sp. nov. can be easily separated from the male of M. longipalpis by the different length of the palpal femur and tibia (cf. Figs. 4A, B View FIGURE 4 vs. 7A, B). The female of M. suzukii sp. nov. is distinguished from the female of other congeners, including M. longipalpis , by the different shape of internal genitalia (e.g., smaller and rounded spermathecae (S) in contrast with larger, sac-like S in M. longipalpis , cf. Figs. 5A, B View FIGURE 5 and 6E View FIGURE 6 vs. Figs. 8E, F View FIGURE 8 ).
Description. Male (holotype). Habitus as in Fig. 5C View FIGURE 5 . Total length: 1.66; prosoma 0.72 long, 0.63 wide. Carapace brownish with sightly lighter central area. Median groove, cervical grooves and radial furrows distinct. Cephalic area poorly defined, slightly raised from carapace. Sternum uniformly brownish. Six eyes all well-developed. ALE=0.05, PLE=0.05, PME=0.05, ALE-PLE=0.01, PLE-PME=0.03. Chelicera, labium and maxillae uniformly brownish. Chelicera bearing a single row of 8 denticles on promargin; denticles on retromargin missing ( Fig. 6D View FIGURE 6 ). Legs uniformly light brown. Leg formula: I, IV, II, III. Leg measurements: I = 4.67 (1.42, 0.22, 1.74, 1.38, 0.91), II = 3.75 (1.06, 0.22, 1.07, 0.81, 0.59), III = 3.09 (0.87, 0.21, 0.80, 0.73, 0.48), IV = 4.03 (1.19, 0.23, 1.26, 1.03, 0.59). Opisthosoma yellowish with faint dorsal marks. Male palp as in Figs. 4A–F View FIGURE 4 , 6A–C View FIGURE 6 . Femur with several long and robust spines on its ventral and dorsal margins. Patella and tibia both elongated, approximately of the same length. Two apophyses (TA) on retrodistal margin of tibia close to each other; ventral apophysis large and lanceolate; dorsal apophysis spine-like, larger at its base and ending with a sharp and long tip ( Figs. 4D, E View FIGURE 4 and 6B View FIGURE 6 ). Cymbium bearing several long and thin spines, ending pointy and with a robust spine at its apex (= tarsal spur). Bulb with two sclerite: median sclerite (MS) spine-like, long and thin; prolateral sclerite (PS) wide and transparent, ribbon-like and wrapped around MS. Embolus (E) long and robust, thread-like and laterally flattened, ending with a rounded tip slightly curved dorsally ( Figs. 4C–F View FIGURE 4 and 6A–C View FIGURE 6 ).
Female (one of the paratypes). Habitus as in Fig. 5D, E View FIGURE 5 . Total length: 1.59; prosoma 0.69 long, 0.63 wide. General coloration and pattern as in male. Frontal view of cephalic area as in Fig. 5E View FIGURE 5 . ALE=0.05, PLE=0.05, PME=0.05, ALE-PLE=0.01, PLE-PME=0.03. Leg formula: I, IV, II, III. Leg measurements: I = 4.69 (1.28, 0.23, 1.42, 1.09, 0.67), II = 3.51 (1.01, 0.21, 0.95, 0.77, 0.57), III = 3.03 (0.85, 0.23, 0.76, 0.69, 0.50), IV = 4.09 (1.12, 0.22, 1.19, 0.95, 0.61). Opisthosoma brownish or yellowish, strongly wrinkled in the frontal part ( Fig. 5E View FIGURE 5 ). Other characters as in male. Internal genitalia as in Fig. 5A, B View FIGURE 5 , 6E View FIGURE 6 . Atrium (AT) wide, cup-shaped. Spermathecae stalk (SS) starting at sides of atrium, first bending inward with a comma course, then reaching spermathecae (S) after one convolution. Spermathecae small and round, separated from each other by two and a half of their diameter.
Size variation: Male (based on 4 specimens): total length: 1.55–1.66, Prosoma 0.65–0.72 long, 0.63–0.66 wide. Female (based on 2 specimens): total length: 1.59–1.70, prosoma 0.69–0.72 long, 0.63–0.66wide.
Distribution. Known only from the type locality ( Fig. 10 View FIGURE 10 ).
Habitat. Caves. The new species was found in empty spaces under stones and in recesses of the floor in the twilight zone of a short and humid cave.
Remarks. M. suzukii sp. nov. shows reduced pigmentation and faint dorsal color pattern. Despite lacking any real troglobitic adaptation, the collecting environment suggests troglophilic habits. However, we do not exclude that this species might also inhabit screes and other shallow subterranean habitats or even external environments (e.g., forest litter). According to Shimojana (1977, pg. 347) Falcileptoneta okinawaensis was also recorded from the same cave where M. suzukii sp. nov. was collected. Despite extensive collections inside the cave, we could not find any other leptonetid species. In addition, the records of F. okinawaensis by Shimojana in Tiragama cave were based on females only. Due to the external similarities of females in Leptonetidae and the lack of information about the internal genitalia of these species, it is possible that such records refer to misidentified samples of M. suzukii sp. nov.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |