
Trididemnum alexi, Lambert, 2003
|
publication ID |
https://doi.org/ 10.5281/zenodo.4525061 |
|
persistent identifier |
https://treatment.plazi.org/id/DB2987AE-FFDC-9129-FCD2-A08A970AC142 |
|
treatment provided by |
Felipe |
|
scientific name |
Trididemnum alexi |
| status |
sp. nov. |
Trididemnum alexi n. sp.
( Figs 1-3 View FIG View FIG )
TYPE MATERIAL. — Holotype: colony collected by C. Lambert on 28.V.2001, from a rope suspended from a floating dock at the town of Friday Harbor, WA, at an approximate depth of 2 m ( MNHN A2 About MNHN TRI 162 ) . Paratype: a second colony 4 cm in greatest length collected on 28.V.2001, from the same rope, growing on a mussel Mytilus sp. ( CASIZ 162519 ) ; companion piece from same colony in 95% ethanol ( CASIZ 162520 ). Additional paratypes collected by C. Lambert from the same rope on 24.VII.2001, 1 colony ( MNHN A2 About MNHN TRI 164 24/VII/2001) and on 29.VIII.2001, 2 colonies ( USNM 1006926 About USNM ) .
OTHER MATERIAL EXAMINED. — USA. Friday Harbor, WA, Town dock, 5. VI.1999, 1 colony (specimen lost).
San Juan Archipelago, Rock Pt, off W side of Lopez I., dredged at 80 m, 3.VIII.1999, D. Duggins coll., 1 small colony growing around the bivalve Mytilimeria nuttalli Conrad, 1837 and heavily parasitized by Botryllophyllus sp., a red copepod with red eggs (identified by S. Ooishi); 2 nearly mature unhatched tadpole larvae found in the colony ( MNHN A2 TRI 163).
Canada. British Columbia, Vancouver I., Canoe Bay Marina, 24.IX.2002, 1 colony, c. 15 cm depth on Mytilus sp. on underside of black floating boat bumper tied to a dock, no larvae, colony surface heavily infiltrated by numerous burrowing amphipods.
British Columbia, Victoria, Fisherman’s Wharf floating docks, 25.IX.2002, 3 colonies, the largest 2 cm thick and 5.5 cm in greatest length, c. 2 m depth on sheet of white plastic tied to long rope, no larvae.
ETYMOLOGY. — It gives me great pleasure to name this species after my grandson, Alexander Ivan Coleman. I hope it will inspire him to pursue a career in biology like his parents and maternal grandparents.
DESCRIPTION
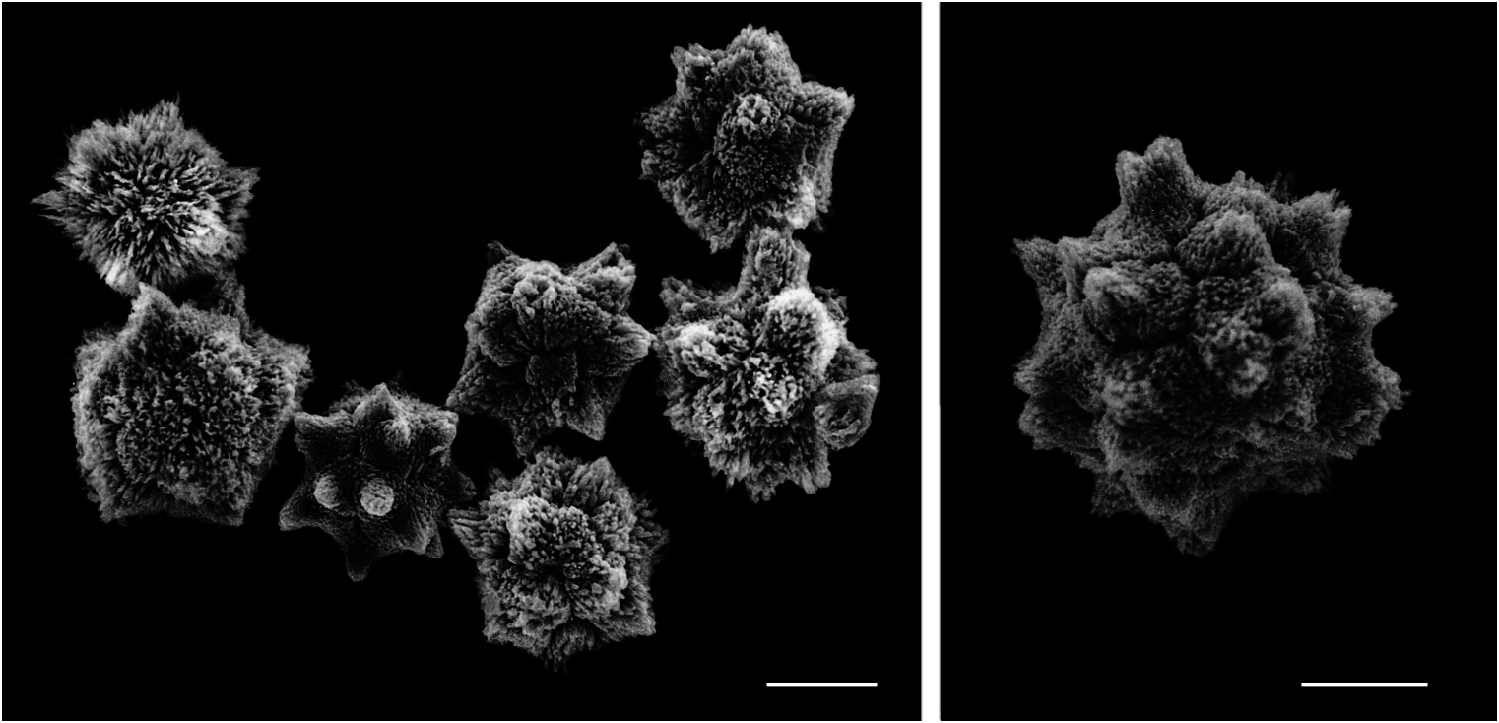
The holotype is 5 cm in greatest length, 1-2 mm thick at the edges but about 2 cm thick in the middle region. The largest paratype is 8 cm in greatest length and also up to 2 cm thick. The structure is complex because the colony has low rounded thickened lobes, is somewhat folded and contains many large ramifying hypozooidal cloacal canals that meander through the tunic. The reddish-brown tunic is smooth, soft and easily torn, with a thick (200-400 µm) superficial blad- der cell layer containing pigment granules but lacking spicules. Different colonies vary somewhat in the intensity of coloration, depending on the concentration of pigment. Under the superficial layer the tunic contains small irregularly stellate spicules ( Fig. 1 View FIG ), 20-30 µm in diameter, with eight to 10 short rays in the equatorial plane. Though the spicules are numerous, they are not densely packed but scattered thinly throughout all layers of the tunic except the bladder cell layer. The pigment and spicules render the tunic opaque so that only the oral openings of the zooids can be seen in an undissected colony. The zooids are located in the upper stratum (above the cloacal canals) and the larvae in the basal zooid-free stratum (below the cloacal canals), with thick columns of tunic here and there connecting the two layers. The zooids do not appear to be in definite systems. Each oral siphon opens separately on the tunic surface. The spicules in the oral lobes are denser in three of the tunic lobes than in the other three. The infrequent common cloacal openings are smooth-edged, large, round and far apart, bordered by thin spicule-free tunic and very little pigment.
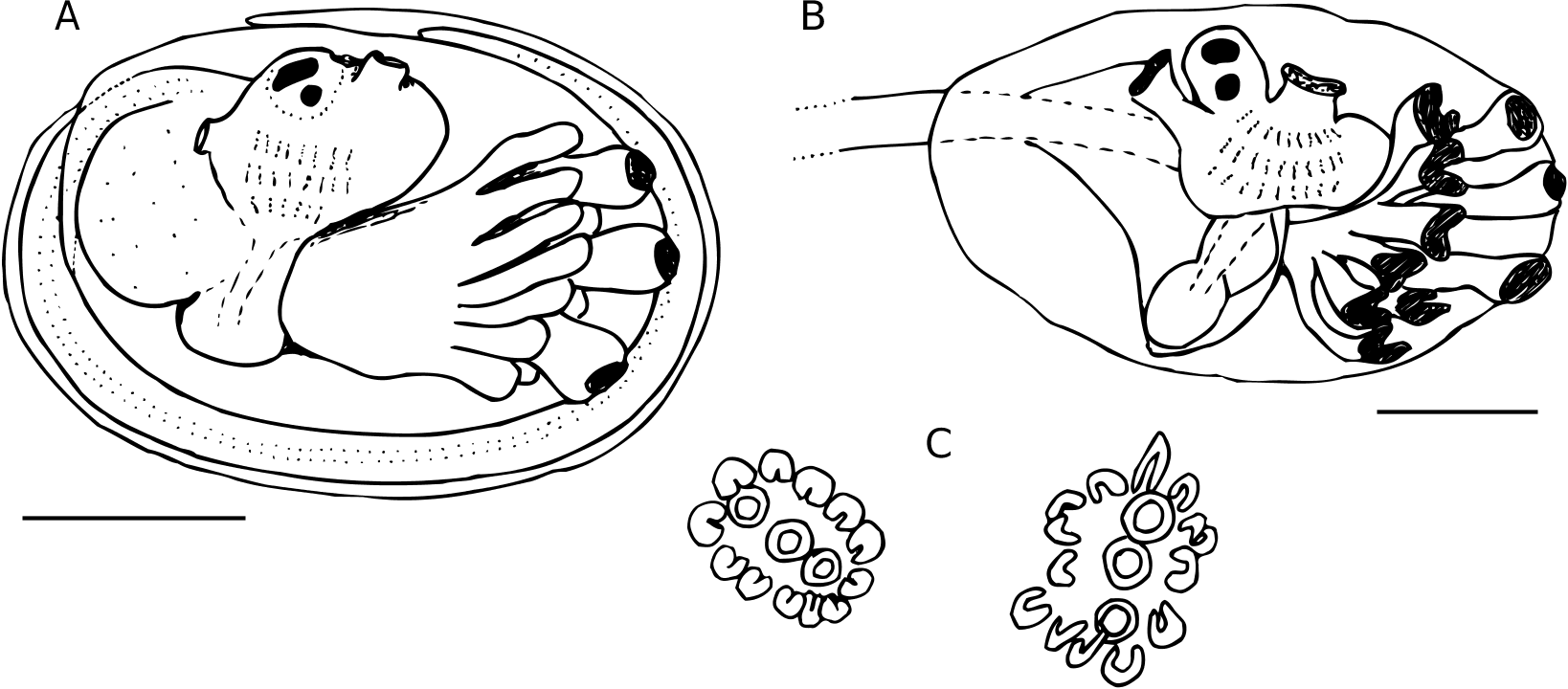
The zooids are about 1.8-2.2 mm long in the preserved state, and heavily pigmented the same reddish brown hue as the tunic. The oral siphon has six short pointed lobes. It was not possible to count the oral tentacles in most zooids, but in one well relaxed individual there were 16 oral tentacles of two sizes, with four longer ones evenly spaced around the base of the oral siphon separating four groups of three shorter tentacles. The three rows of stigmata are elongate, with a large unperforated area just posterior to the peribranchial region (Fig. 2A, B). There are usually 12-13 stigmata per side in row 1, 12 in row 2, and 10- 12 in row 3. Two backward-curved dorsal languets are present. The unlobed atrial siphon, large and tubular or slightly flaring, is directed dorsally or posteriorly, and is located at the level of the middle row of stigmata. It opens into the large hypozooidal common cloacal cavity. The spicule-forming lateral organs of the thorax ( Kniprath & Lafargue 1980) are large, densely filled with very small spicules, and located between the second and third rows of stigmata or over the third row (Fig. 2B, dashed circle).
The esophagus is very long and may be constrict- ed by a blood vessel or ectodermal appendage that wraps around the zooid at this point and extends out into the tunic (Fig. 2A). The retractor process is present in only some of the zooids and may be very short or almost as long as the esophagus; it originates from the base of the thorax alongside the top ventral side of the esophagus (Fig. 2B). The stomach is large, globular, and smooth. A single large egg is present in most zooids. The sperm duct is a flattened cap over the single testis, with eight or nine coils in most zooids; a few have seven and two were observed to have 10. Many of the zooids are forming a bud: the adult thorax forms a new abdomen, while the adult abdomen forms a new thorax (Fig. 2C).
In the basal layer of tunic are the developing embryos ( Fig. 3A, B View FIG ), some in an advanced stage apparently ready to be released. The embryos do not appear to be connected to any zooids. The fully developed unhatched tadpoles are 0.8-1.0 mm in trunk length, with three adhesive papillae and in most cases 14 long stout ampullae, seven on each side. Rarely there is a 15 th median ampulla anteri- or to the papillae. Both ocellus and otolith are present, and three faint rows of stigmata can be seen in the developing branchial sac. A hatched tadpole that was still in the cloacal canal and a fully mature unhatched tadpole had a trunk length of 1 mm. In these two largest tadpoles the lateral ampullae are not circular and cylindrical at their tips but are curved ( Fig. 3B, C View FIG ), similar to those described and illustrated by Sanamyan (1999) for T. tenerum ( Verrill, 1871) .
REMARKS
The only other species of Trididemnum known from the San Juan Archipelago is T. opacum ( Ritter, 1907) , a common species that is more thinly encrusting and grey or a pale flesh color when alive. It has much denser stellate spicules throughout the tunic with longer points than in the new species, the atrial siphon is posterior to the third row of stigmata, and there are only about six coils of the sperm duct ( Table 1). It is possible that T. alexi n. sp. is the same species as Huntsman’s (1912a) Trididemnum sp. A from the east coast of Vancouver Island in the Departure Bay area; he wrote that “a very dark Trididemnum with few or occasionally no (?) [his question mark] spicules was taken several times in the dredge”. This specimen has apparently been lost, as persistent searching of several museums has yielded no trace of his specimen.
The new species resembles Trididemnum strangulatum ( Ritter, 1901) from Alaska in its overall shape and the long constricted esophagus, a character present in most Trididemnum species and thus considered by Van Name (1945) of no taxonomic importance. However, Ritter described T. strangulatum as translucent and “ashen” and with only four coils of the sperm duct. He did not measure the spicules, stating only that they were “short and blunt”. Van Name (1945) identified as T. strangulatum a colony from Chignik Bay, Alaska, with 20 µm spicules but did not include other morphological details; he did not examine Ritter’s specimens. Van Name indicated that his specimen is in the USNM, but it could not be located to examine for this study, nor could Ritter’s type specimen. Sanamyan (1999) tentatively identified as T. strangulatum two colonies from Sanak Island, Gulf of Alaska; however in his specimens the spicules are large (up to 50 µm) with “prominent conical rays” and twice as many coils of the sperm duct as in Ritter’s specimens. Trididemnum tenerum ( Verrill, 1871) is considered a circumbo- real species occurring in both the north Atlantic and north Pacific. Van Name (1945) reexamined Verrill’s specimens and gives a detailed description. Verrill described living colonies as translucent and did not mention any color. The zooids apparently resemble T. opacum and T. alexi n. sp. morphologically but no accurate description of the spicules is available. Van Name (1945) described the holotype as having needle-like radiating crystals; this is surely a sign of the original spicules having dissolved in the preservative and then the calcium carbonate recrystallized in a needle-like pattern. This is not an unusual phenomenon in didemnids if the preservative is not properly buffered, and happened to some of my own samples occasionally in years past. I have never seen this type of spicule in fresh living colonies, only in fixed material. Sanamyan (1999) tentatively identified a colony from Kamchatka as T. aff. tenerum , with large (50+ µm) spicules with rounded rays and also spicules with needle-like rays; the latter may have formed by dissolution and recrystallization within the spicular envelopes. The larvae are much larger than T. alexi larvae and have a total of only eight or nine lateral ampullae.
Thus Trididemnum alexi n. sp. is distinguished from all known NE Pacific Trididemnum species by the following constellation of characters, as listed in Table 1: smooth tunic and zooids dark reddish brown with abundant pigment granules; colony up to 2 cm thick with large ramifying hypozooidal canals and basal portion containing the larvae; irregularly stellate spicules 20-30 µm in diameter with short pointed rays (eight to 10 in the equatorial plane) thinly scattered through the tunic but absent from the 200-400 µm thick superficial bladder cell layer; thorax with 12-13 stigmata per half row in the first two rows on average and one or two fewer in the third row; atrial siphon at the level of the second row of stigmata; single testis covered by the sperm duct with eight or nine coils; larvae 0.8-1.0 mm in trunk length with 13-15 (usually 14) lateral ampullae with curved tips when fully mature.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.