
Daurlong wangi, Wang & Cau & Guo & Ma & Qing & Liu, 2022
|
publication ID |
https://doi.org/ 10.1038/s41598-022-24602-x |
|
persistent identifier |
https://treatment.plazi.org/id/DC228C6D-092A-A041-FB66-FA1E26FCF9B8 |
|
treatment provided by |
Felipe |
|
scientific name |
Daurlong wangi |
| status |
gen. et sp. nov. |
Daurlong wangi gen. et sp. nov.
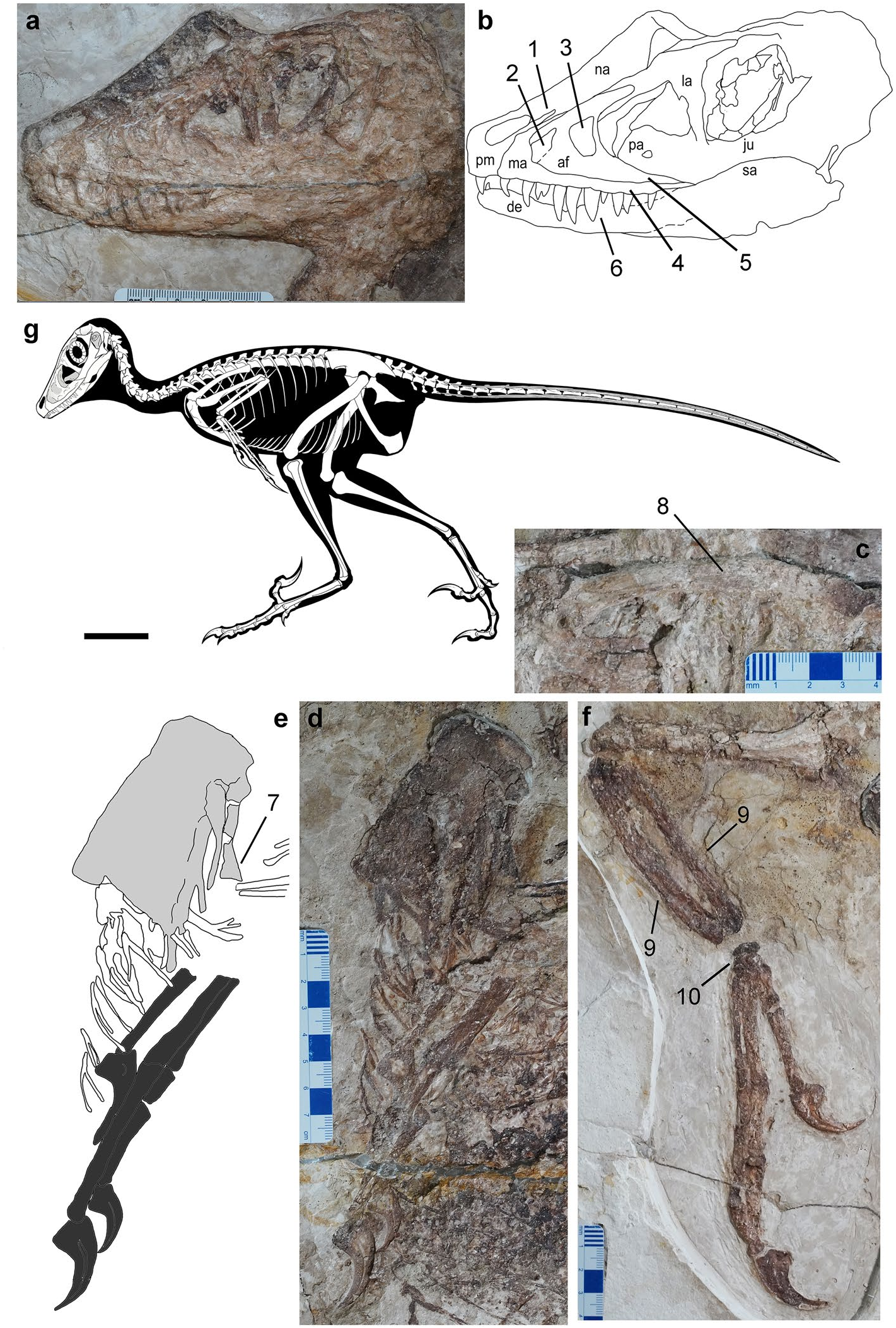
Holotype IMMNH-PV00731 , an almost complete dromaeosaurid ( Figs. 1 View Figure 1 , 2 View Figure 2 , Supplementary information). Locality and Horizon Pigeon Hill , Morin Dawa Daur Autonomous Banner, Inner Mongolia Autonomous Region (N 48°39′40.76″ / E 123°52′ 41.15″); Longjiang Formation, Lower Cretaceous. GoogleMaps
Etymology The genus name is derived from the Daur Nation, and the Chinese AE ("lóng") for "dragon". The species name honors Mr. Wang Junyou, director of the IMMNH.
Diagnosis Mid-sized dromaeosaurid with (autapomorphies marked by asterisk): slender subnarial ramus of premaxilla extended caudally well beyond the external naris; large, trapezoid promaxillary recess placed at the rostroventral corner of antorbital fossa*; maxillary fossa large, shallow and caudodorsally located, so that the pila promaxillaris is wider than the pila interfenestralis *; stepped transition from the subcutaneous surface of maxillary ventral ramus to the antorbital fossa; fan-shaped distal end of first sternal rib*. Differential diagnosis: Daurlong further differs from Tianyuraptor because it bears longer and more robust maxillary teeth and a more robust ulna. Daurlong further differs from Zhenyuanlong because it lacks a pitted ventral ramus of the antorbital fossa, lacks markedly concave distal margins in maxillary tooth crowns, bears a bowed scapula, a more robust radius, and a wider overlap of the semilunate carpal over metacarpal II ( Fig. 3 View Figure 3 ).
Description and comparison. The holotype of Daurlong wangi is an almost complete and articulated skeleton with a length of about 150 cm ( Fig. 1 View Figure 1 ). Part of the ribcage and the corresponding gastral region are overlapped by the lef forelimb (segments of the humerus and forearm are missing) and suffered some crushing during diagenesis. The specimen is 85% the size of Tianyuraptor ostromi holotype 6, 93% the size of Zhenyuanlong suni holotype 7, and between 115 and 350% the size of the Jehol Biota microraptorines 5, 8. The skull is almost perfectly articulated, except for the missing nasal ramus of the lef premaxilla and the partially displaced dorsal parts of the right nasal and lacrimal ( Figs. 1 View Figure 1 , 3 View Figure 3 ). A similar displacement of nasal and lacrimal is visible in the holotype of Zhenyuanlong 7. The skull is about 94% of femur length, comparable to most taxa (e.g., Halszkaraptor , Microraptor and Saurornitholestes ) whose skulls are approximately 92–95% of femur length 8 – 13. In Zhenyuanlong , the skull is about 86% of femur length 7, and in Tianyuraptor the skull is longer than the femur 6. In Bambiraptor and Sinornithosaurus , the skull is about 103–105% longer than the femur 11. In Velociraptor , the skull is between 117 and 128% of femur length, due to the relatively elongate snout 10. The rostral margin of the premaxilla forms a right angle with the occlusal margin, similar to Velociraptor , Zhenyuanlong and Saurornitholestes 2, 10, 12, differing from the shallower premaxillae with acute rostroventral corner in Microraptor and Sinornithosaurus 4, 11 and the shallow platyrostral premaxilla of Halszkaraptor 13. The subnarial process of the premaxilla extends caudodorsally well beyond the caudal margin of the external naris, as in Velociraptor and Linheraptor 12, 14, differing from the relatively shorter processes in Microraptor and Zhenyuanlong 2, 5. Saurornitholestes shows an intermediate condition. The external naris is relatively small, tear-shaped with the long axis directed caudodorsally, similar to Zhenyuanlong 7 and differing from the relatively larger external naris in Microraptor 4. In eudromaeosaurs, the external naris is relatively shorter and its long axis is subparallel to the oral margin 10, 12.
The subcutaneous part of the rostral ramus of the maxilla is relatively small and triangular, taller than long and similar to Zhenyuanlong 7 and Wulong 15, differing from the relatively more elongate ramus in Halszkaraptor 13, Microraptor 4 and in some eudromaeosaurs 14, 16, 17. The antorbital fossa is large, longer than tall and covering more than two-thirds of the maxilla, similar to Zhenyuanlong and Wulong 7, 15, differing from the relatively shorter excavation seen in some eudromaeosaurs (e.g., Acheroraptor 17). As in Zhenyuanlong , the rostral margin of the antorbital fossa is at the level of the second maxillary tooth, more rostral than in other dromaeosaurids 4, 7, 12. The promaxillary fenestra is relatively large, as in other Jehol Biota dromaeosaurids 6 and is placed adjacent to the rostroventral corner of the antorbital fossa, differing from the more caudally placed fenestra in Sinornithosaurus and Zhenyuanlong 7, 11. In Microraptor , the promaxillary fenestra is narrower rostrocaudally and elongated dorsoventrally 4. The promaxillary fenestra is trapezoid in lateral view, with straight rostral, caudal and ventral margins, and a caudodorsally slanted dorsal margin. The maxillary recess is a large shallow fossa delimited by curved ridges paralleling the rostral margin of the antorbital fenestra. The pila promaxillaris is rostrocaudally wider than the pila interfenestralis (Supplementary information), a condition differentiating Daurlong from both Tianyuraptor and Zhenyuanlong , which show a relatively narrower pila promaxillaris 6, 7. In microraptorines 4, 11 the area around the pila promaxillaris and the region underneath the antorbital fenestra are excavated by a series of small pits and ridges, absent in Daurlong . The caudoventral ramus of the antorbital fossa lacks the pits and crests present in Zhenyuanlong 7. The dorsoventrally shallow subcutaneous surface of the ventral ramus is separated from the antorbital fossa by a stepped margin, differing from Zhenyuanlong which shows a distinct antorbital rim 7. As in other dromaeosaurids 17 – 19, the postrantral wall is widely exposed in lateral view.
The lacrimal is T-shaped as in most dromaeosaurids 4, 10 – 12. The frontal widely contributes to the dorsal margin of the orbit. The orbit is ovoid-shaped with the long axis directed rostroventrally-caudodorsally ( Fig. 1 View Figure 1 ). The scleral ring is nearly completely preserved inside the orbit. Its dorsoventral axis coincides with that of the orbital fenestra, and does not appear significantly deformed, allowing for an accurate measurement of its inner and external diameters. The scleral ring is large (external diameter about 93% of orbital diameter) and moderately slender. The rostral ramus of the postorbital is upturned, forming an obtuse angle with the caudal ramus, which is instead perpendicular to the ventral ramus. The elements of the caudal portion of the skull are badly preserved. The dentary shows a length/depth ratio of approximately 8, proportionally intermediate between the more gracile conditions in Microraptor and Velociraptor , and the relatively stouter dentary of Saurornitholestes 4, 10, 12. The dorsal margin is slanted rostroventrally at its rostralmost end, then, caudal to the second alveolus, it is parallel to most of the ventral margin, as in most non-dromaeosaurine dromaeosaurids 4, 10. The ventral margin of the dentary gradually tapers rostrodorsally from the rostral sixth of the bone, and then is inclined caudodorsally along the caudal third. The postdentary elements of the mandible are badly preserved.
The single premaxillary tooth crown preserved is unserrated, bears a sharp apex, and is slightly curved distally near the apex. Ten teeth are preserved in the maxilla, with an additional tooth missing. The crowns are more robust and elongate than those of Tianyuraptor 6. The middle maxillary teeth are the longest and fang-like, similar to those of Microraptor 4. The crowns of the second, fourth and last preserved maxillary tooth bear dense serrations along their distal carinae. All maxillary teeth are blade-shaped and only slightly curved distally near the apex, differing from the more curved teeth of Zhenyuanlong 7. Six dentary teeth can be recognized. They are much smaller than the maxillary teeth. Ten cervical vertebrae are preserved in articulation, but badly preserved ( Fig. 2 View Figure 2 ). The cranial neural spines of the dorsal series are rectangular and strongly inclined caudodorsally. The caudal neural spines of the dorsal series are square-shaped, with their long axis oriented dorsally, almost vertical to the corresponding centra ( Fig. 2 View Figure 2 ). The sacrum includes six vertebrae. The sutures along the sacral centra are obliterated, except for a faint suture visible between centra 2 and 3. The first caudal vertebra is similar in size to the last sacral vertebra. The tail is complete, approximately 4.4 times longer than the femur, but the number of the vertebrae is uncertain ( Fig. 2 View Figure 2 ). Most of the caudal vertebrae are encased in the caudotheca 21 as in microraptorines andeudromaeosaurs 4, 20. Daurlong is similar to other Jehol Biota dromaeosaurids 4, 6, 7 in having the caudotheca extended through the rostralmost vertebrae, differing from eudromaeosaurs where the bony rods bundle begins distal to caudal 620, 21. The scapula is strap-like and uniformly bowed dorsally, differing from the straight scapula of Zhenyuanlong 7. As in most paravians, the scapula is shorter than the humerus and more gracile than midshaf diameter of the ulna, with no distal expansion. The lef coracoid is partially preserved. The bone is fused to the scapula, and both contribute to the laterally-facing glenoid, as in other paravians 1, 22. There is no evidence of the large coracoid fenestra present in some microraptorines 1, 4. The sternum is large and subrectangular with the long axis as long as the humeral shaf ( Fig. 2 View Figure 2 ). It appears as a single unpaired element, as in Microraptor 21, and differing from the incompletely fused elements in Halszkaraptor and eudromaeosaurs 1, 14, 24. Four pairs of sternal ribs articulate with the sternum, as in Microraptor and Sinornithosaurus , differing from Zhongjianosaurus which shows five pairs 25. The first pair of sternal ribs is more robust than the others and shows an elongate fanshaped morphology expanded laterodistally, differing from the slender shaf and the spoon-shaped distal end shared by other dromaeosaurids 5, 25. In some avialans 26, the caudalmost sternal rib is the most robust and shows an expanded distal end similar to the condition in Daurlong . As in other paravians, the gastralia contact the pubis shaf in a position more proximal than in other theropods, where instead such contact is usually placed on the pubic foot 27.
The forelimb length is less than 60% of the hindlimb. Among dromaeosaurids, comparably short forelimbs are shared with Austroraptor 19,halszkaraptorines 9, Tianyuraptor 6 and Zhenyuanlong 7, and differ from the relatively longer forelimbs in Buitreraptor 28,microraptorines 4, 5, and some eudromaeosaurs 20. The deltopectoral crest is not prominently developed. The ulna is slightly bowed caudally and more robust than in Tianyuraptor 6. The radial shaf is about half the thickness of the ulna and nearly equal to the mid-shaf of the manual phalanx I-1: in Zhenyuanlong the radius is apomorphically more gracile 7. In the carpus, the stout scapholunare articulates distally with the semilunate carpal, which, in turn, overlaps the whole extent of the proximal end of metacarpal II, differing from the more limited overlap in Zhenyuanlong 7. The combined metacarpal I + manual phalanx I-1 complex is longer than metacarpal II, differing from microraptorines which have metacarpal II longer than metacarpal I + phalanx I- 14, 5. Manual digit II is the longest and bears the largest claw. As in microraptorines and eudromaeosaurs 4, 5, 20 manual phalanx III-2 is shortened, being about half the length of phalanx III-1. All the three recurved manual claws bear prominent flexor tubercles and horny sheaths. The ilium is 64% of the femoral length. The preacetabular process is longer than the postacetabular process of the ilium, a feature shared with Halszkaraptor , Tianyuraptor and Unenlagia among dromaeosaurids 6, 9, 29, and comparable to Rahonavis and most avialans 30. As in other dromaeosaurids 5, the postacetabular process is gradually declined caudoventrally but does not extend below the level of the ischial peduncle. The pubic shaf is straight and caudoventrally oriented. The pubic foot is broadly rounded and dorsoventrally expanded, as in other Jehol Biota dromaeosaurids 2. The ischium is about half the pubis in length, lacks dorsal processes along the straight dorsal margin and ends distally in a poorly curved tip. The obturator process is triangular, as long as deep, and placed at mid-lenght of the ventral margin, differing from the prominent and more distally-placed process in microraptorines 5.
The femur is 91% of the tibia in length (in both Tianyuraptor and Zhenyuanlong , the femur is <77% of the tibia). In theropods, the femur-to-tibia ratio is allometrically controlled 31, and results higher in large-bodied dromaeosaurids than in small-bodied taxa 32. The feet are incomplete, with the medialmost elements (i.e., toes I and II) badly preserved and overlapped by other foot bones. Metatarsal III and IV are nearly equal in length and about 61% of the femoral length. The preserved four pedal claws are comparable in size, are strongly recurved and bear low but prominent flexor tubercles.
The plumage is preserved along the dorsal margin of the postorbital part of the skull, adjacent to the presacral neural spines ( Fig. 1d View Figure 1 ) and along the edges of the tail. No feathers are preserved close to the limbs or along the ventral margin of the body: it is unclear if long pennaceous remiges and rectrices were present as in Zhenyuanlong 7. The plumage along the dorsal margin of the pre-caudal part of the skeleton is preserved as a series of compound structures containing several filaments joined at their proximal ends, similar to the condition in Sinornithosaurus 33. Short pennaceous feathers are preserved along most of the margin of the caudotheca. The tail feathers appear symmetrical, oriented backward and forming a low angle (~ 15–20°) with the proximodistal axis of the adjacent tail vertebrae. Scanning electron microscopy (SEM) of sampled portions of the tegumentary remains failed to identify melanosomes 34 – 36. The caudal half of the abdominal cavity of the theropod is occupied by a black-bluish layer which is bound by the gastral basket (ventrally) and the pubis (caudally) ( Figs. 1 View Figure 1 , 2 View Figure 2 , Supplementary Information). The rostral margin of the layer is placed ventral to the 9th dorsal centrum, where it extends dorsoventrally along the dorsal half of the abdominal cavity. The black-bluish layer reaches its maximum depth between the 10th and the 11th dorsal vertebrae, where it reaches the ventralmost part of the gastral basket. Caudal to the 11th dorsal vertebra, the layer is limited to the dorsal end of the abdominal cavity.
The imprint of the partial skeleton of an anuran is preserved in the same slab of IMMNH-PV00731, adjacent to the theropod metatarsi ( Fig. 1 View Figure 1 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.