
Gonioinfradens giardi ( Nobili, 1905 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4370.5.9 |
|
publication LSID |
lsid:zoobank.org:pub:29848543-5479-4280-8F31-6E1FAD2398DB |
|
DOI |
https://doi.org/10.5281/zenodo.5980610 |
|
persistent identifier |
https://treatment.plazi.org/id/DC2D878A-FFB2-E720-4EFF-F747FEC5F88A |
|
treatment provided by |
Plazi (2018-01-15 06:40:59, last updated 2024-11-28 09:00:11) |
|
scientific name |
Gonioinfradens giardi ( Nobili, 1905 ) |
| status |
|
Gonioinfradens giardi ( Nobili, 1905) View in CoL
( Figs 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Thalamita Giardi Nobili, 1905: 164 View in CoL .
Charybdis (Goniosoma) Giardi. View in CoL — Nobili 1906: 115, pl. 5 fig. 23, pl. 7 fig. 34.
Charybdis giardi View in CoL . — Balss 1924: 3.
Charybdis (Gonioinfradens) paucidentata View in CoL .— Leene 1938: 131, figs 74‒76.— Stephensen 1945: 119.
Gonioinfradens paucidentata View in CoL . — Apel & Spiridonov 1998: 223, figs 40‒41, pl. 7.— Apel 2001: 72.— Naderloo & Sari 2007: 343.
Gonioinfradens paucidentatus View in CoL . — Corsini-Foka et al. 2010: 331, fig. 2.— Karhan & Yokeş 2012: 118, fig. 2.
Type material. Gonioinfradens giardi . Lectotype: Persian Gulf, United Arab Emirates, Mission J. Bonnier and Ch. Pérez, Stn XLVII, pearl oysters banc, 25°10’N 55°10’E to 24°55’N 54°40’E, 10–15 fathoms, 1901, M 42.7 mm ( MNHN IU 2014 11969 , previously MNHN B.5657) id. as Thalamita giardi . GoogleMaps Paralectotypes: same data as lectotype, F 38.3 mm ( MNHN B.5657 ) ; Stn LII, pearl oysters banc, SW of Arzana I., SE of Zirkuh I., M 30.6 mm, F broken ( MNHN B.5659 ) ; Station LIII, NE of Arzana I., F (broken) 27.8 mm, F immature 10.3 mm ( MNHN B.5656 ) ; same data, 2 immature 9.9, 6.6 mm ( MNHN B.5658 ) .
Other material examined. Djibouti, Rècif d’Ambouli, Mission J.L. Dantan, 1933, M immature 19.0 mm ( MNHN) .
Israel, Ashdod , port breakwater, 31°50.26’N, 34°37.93’E, 5–15 m, 14 May 2017, coll. R. Gevili, M 46.5 mm (SMNHTAU-AR 29776). GoogleMaps
Description. Carapace 1.35–1.40 times as broad as long in adult males, regions ill-defined, smooth, glabrous. Faint, minutely granular transverse lines anteriorly: a pair each of frontal, protogastric, metagastric, epibranchial and a nearly straight unbroken line on mesogastric; frontal and mesogastric lines barely noticeable, only epibranchial line raised. Front, wider than posterior edge, cut into six lobes: median lobes rounded, submedian lobes slightly less prominent, skewed distad, anterior margins on slightly higher plane than latter, lateral lobes with oblique mesial margin and rounded lateral margin, projecting forward to the level of median lobes, separated from submedian lobes by wide, V-shaped incision. Inner supra-orbital angles raised higher than frontal lobes, larger, but similar in shape to submedian lobes. Basal antennal segment broad, roughly granular, bearing medially single prominent spine, curved distad; antennal flagellum excluded from orbital hiatus. Orbits wide, upper orbital margin bifissured, lower margin unifissured; inner infra-orbital angle prominent, triangular, tooth-like. Sub-orbital, subhepatic and pterygostomian regions sparsely pilose. Anterolateral border bearing four well-developed, finely granular teeth with anterior margin concave, posterior margin convex. Small ancillary teeth in between larger teeth: pointed triangular tooth basally on posterior margin of anterior tooth, and successively smaller, obtuse teeth in between large teeth. Posteriormost tooth markedly smaller. Posterolateral margins oblique, converging. Posterior margin nearly horizontal.
Sternum bare and smooth, save for sparse punctae.
Chelipeds unequal, massive. Merus bearing three spines on anterior margin, proximalmost smallest, none on posterior margin. Carpus bearing large spine on inner angle and three smaller spines on outer face, arranged equilaterally. Manus bearing spine proximally near carpal articulation, two closely set spines medially on upper surface ( Fig. 3A–D View FIGURE 3 ), proximalmost spine pointing mesially, swollen above articulation with moveable finger; lateral margins smooth, but for low rounded protuberance distally near base of moveable finger; inner lower margin with closely set granulation proximally. Fingers deeply grooved, their cutting edges lined with rounded tubercules of unequal size, proximally with molariform tooth in larger chela.
Ambulatory legs laterally compressed, propodus and dactylus 1–3 grooved laterally. Merus of natatory legs bearing prominent spine subdistally on posterior margin, dactylus and propodus foliaceous, propodus with a row of spinules on posterior margin.
Male abdomen triangular, segments 3–5 fused, penultimate segment broader than long, lateral margins convex ( Fig. 4A–D View FIGURE 4 ), telson triangular.
G1 bent distad, distally cornute; outer margin lined with long spinules, mesial and ventral margins with rows of short spinules next to the tip ( Fig. 5A View FIGURE 5 ). Female sternopleonal cavity with vulvae on sternite 6, round, slightly hooded ( Fig. 6 View FIGURE 6 ).
Distribution. Red Sea, Gulf of Aden, Gulf of Oman, Arabian Gulf. Recently recorded in the Mediterranean Sea from Turkey and Greece, newly recorded from Israel.
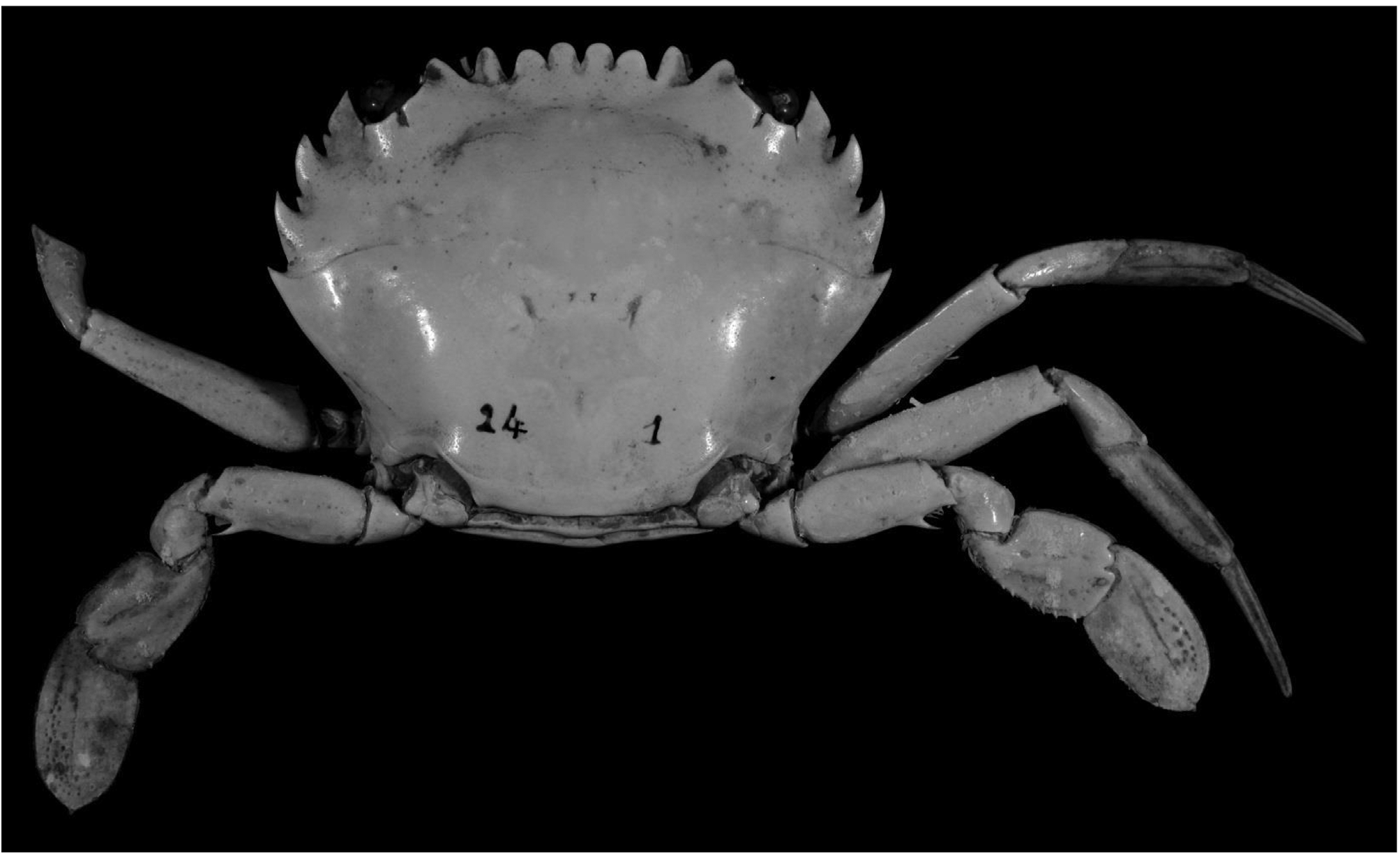
Coloration. The carapace of the specimen collected off the Israeli coastline is mottled reddish brown dorsally, with a distinct Y-shaped dark brown mark centrally. The chelipeds are deep reddish brown with dark tipped fingers; the ambulatory legs reddish with pale colored bands near joints. Spines on the anterolateral margin of the carapace and chelipeds are distally light-banded and dark-tipped ( Fig. 2 View FIGURE 2 ). It is similar in color pattern to specimens from the Arabian Gulf ( Apel & Spridonov 1998: pl. 7.7) and from Rhodes I. ( Corsini-Foka et al. 2010: fig. 2).
Remarks. Nobili’s (1905) brief description of Thalamita Giardi was based on a specimen with carapace length (cl) of 46 mm and width (cw) of 60 mm, of unspecified gender. Subsequently ( Nobili 1906) renamed it Charybdis (Goniosoma) Giardi and listed 9 specimens (2 M in Stn 47, 1 M 2 F in Stn 52, 1 M, 3 F, 2 damaged, in Stn 53), but based his description and figures on the larger male specimen from Stn 53 (cl 42 mm, cw 61 mm). Leene (1938: 131) examined “two cotypes of Charybdis Giardi Nobili (Mission J. Bonnier et Ch. Perez 1901, stat. 52) and two other specimens of the latter species (1901, stat. 53). The cotypes (M and juv.) were much damaged; the larger specimen (F) of stat. 53 is better preserved, the other specimen was juvenile.” Leene’s description and illustration is based on “the largest specimen of stat. 53 of the Ch. Giardi specimens, at the same time comparing it with the original description by G. Nobili and with the cotypes” ( Leene 1938: 131). The material from Stn 47 went unmentioned. Apel & Spiridonov (1998: 224) listed two immature syntype specimens (MNHN B.5658), a male and an immature female (MNHN B.5659), and a female and immature specimen collected NE of Arzanah I. (MNHN B.5657). A recent examination of the material revealed a mismatch in the number of specimens and station numbers with these accounts: the jar labeled as Stn 47 (MNHN B.5657) contained adult male and female specimens, the one labeled as Stn 52 (MNHN B.5659) adult male and single damaged female, and in the two jars labeled Stn 53 – one contained an adult female and an immature female (MNHN B.5656) and the other two immature specimens (MNHN B.5658) – for a total of eight specimens. It seems that over the past century the specimens may have been shifted from one jar to another – the larger male specimen (cl 42 mm) described from Stn 53 by Nobili (1906) is currently in a jar labelled Stn 47 (MNHN B.5657).
As Nobili had not designated a holotype specimen, and some confusion arose since concerning the number of specimens and their collection numbers, all being syntypes, we hereby designate as lectotype the male specimen (cl 42.7 mm), collected in Stn XLVII, pearl oysters banc, Mission J. Bonnier and Ch. Pérez, 1901 (MNHN IU-2014- 11969, previously MNHN B.5657), and the remaining syntypes become paralectotypes.
The female type specimen of Goniosoma paucidentatum A. Milne Edwards , missing its chelae is preserved dry (MNHN 780) ( Fig. 6 View FIGURE 6 ). It accords well with Milne Edwards’ (1861) description. Nobili (1906: 119) considered it very close to Ch. (Goniosoma) Giardi : “ Goniosoma paucidentatum A. Edw. parait bien voisin de cette espèce, mais ses chélipèdes ne sont pas connus et l’article basilaire de l’antenne semble être conformé différemment”. Leene (1938: 131) then erroneously “suppose[d] this species [Ch. Giardi] is identical with G. paucidentatum A. M. Edw. ”, and placed it in synonymy to Ch. (Gonioinfradens) paucidentata . This was accepted by subsequent authors and Ch. (Goniosoma) giardi slid into obscurity despite consistent and obvious differences in color pattern and morphological characters (e.g., Sakai 1976: pl. 130, fig. 1; Poupin 1994: pl. 3e).
Gonioinfradens paucidentata differs from G. giardi in the shape of frontal lobes (Figs 2,7), having the two spines medially on upper surface of palm set further apart ( Fig. 3A, C, E View FIGURE 3 ), short rounded carina distally on inner and outer lateral palmar surfaces ( Fig. 3B, D, F View FIGURE 3 ), lateral margins of penultimate segment of male abdomen nearly parallel, converging distally ( Fig 4A–F View FIGURE 4 ), shape of G1 ( Fig. 5 View FIGURE 5 ) and color pattern.
Based on morphological characters observed in the examined specimens, including type material, and the results of the molecular analyses based on the mitochondrial barcoding gene cytochrome oxidase sub unit I, G. giardi is reinstated as a valid species, and is thus withdrawn from synonymy with G. paucidentata . The former is confined to the Red Sea, Gulf of Aden, Gulf of Oman, Arabian Gulf, and was recently introduced to the Mediterranean through the Suez Canal, whereas the latter is seemingly widely distributed in the Indo-Pacific, recorded from Mauritius, Seychelles and Réunion to Japan, New Caledonia and French Polynesia.
Validated identification is of major concern for management of marine non-indigenous species as taxonomic identity provides information as to possible origin and pathway of introduction ( Ojaveer et al. 2014). We join Evans and Schembri (2017: 199) in calling “…authors and reviewers to exercise caution and check identification carefully”.
Apel, M. & Spiridonov, V. A. (1998) Taxonomy and Zoogeography of the portunid crabs (Crustacea: Decapoda: Brachyura: Portunidae) of the Arabian Gulf and adjacent waters. Fauna of Arabia, 17, 159 - 331.
Apel, M. (2001) Taxonomie und Zoogeographie der Brachyura, Paguidea und Porcellanidae (Crustacea: Decapoda) des Persisch-Arabischen Golfes. Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften. Vorgelegt beim Fachbereich Biologie der Johann Wolfgang Goethe-Universitat in Frankfurt am Main, 255 pp.
Balss, H. (1924) Decapoden des Roten Meeres. III. Die Parthenopiden, Cyclo- und Catometopen. In: Expedition S. M. Schiff Pola in das Rote Meer, nordliche und sudliche Halfte 1895 / 96 - 1897 / 98. Zoologische Ergebnisse. XXXIV. Denkschriften der kaiserlichen Akademie der Wissenschaften zu Wien, Mathematisch-naturwissenschaftliche Klasse, 99 (6), 1 - 18.
Corsini-Foka, M., Pancucci-Papadopoulo, A., Kondilatos, G. & Kalogirou, S. (2010) Gonioinfradens paucidentatus (A. Milne Edwards, 1861) (Crustacea, Decapoda, Portunidae): a new alien crab in the Mediterranean Sea. Mediterranean Marine Science, 11 / 2, 331 - 340. https: // doi. org / 10.12681 / mms. 80
Evans, J. & Schembri, P. J. (2017) On the occurrence of Cephalopholis hemistiktos and C. taeniops (Actinopterygii, Perciformes, Serranidae) in Malta, with corrections of previous misidentifications. Acta Ichthyologica et Piscatoria, 47 (2), 197 - 200. https: // doi. org / 10.3750 / AIEP / 02064
Karhan, S. U. & Yokes, M. B. (2012) An earlier record of the Indo-Pacific swimming crab, Gonioinfradens paucidentatus (A. Milne-Edwards, 1861) (Decapoda, Brachyura, Portunidae) off the Mediterranean coast of Turkey. Crustaceana, 85 (1), 117 - 121. https: // doi. org / 10.1163 / 156854012 X 623575
Leene, J. E. (1938) The Decapoda Brachyura of the Siboga-Expedition. VII. Brachygnatha: Portunidae. Siboga Expeditie Monografie, 39 C 3 (131), i - iv + 1 - 156.
Milne Edwards, A. (1861) Etudes zoologiques sur les Crustaces recents de la famille des Portuniens. Archives du Museum national d'Histoire naturelle, Paris, 10, 309 - 421. https: // doi. org / 10.5962 / bhl. title. 10629
Naderloo, R. & Sari, A. (2007) Subtidal crabs of the Iranian coast of the Persian Gulf. New collections and biogeographic considerations. Aquatic Ecosystem Health and Management, 10 (3), 341 - 349. https: // doi. org / 10.1080 / 14634980701514620
Nobili, M. G. (1905) Decapodes nouveaux des cotes d'Arabie et du Golfe Persique. (Diagnoses preliminaires.) Bulletin du Museum national d'Histoire naturelle, Paris, 11 (3), 158 - 164.
Nobili, M. G. (1906) Mission J. Bonnier et Ch. Perez (Golfe Persique 1901). Crustaces Decapodes et Stomatopodes. Bulletin scientifique de la France et de la Belgique, 40, 13 - 159.
Ojaveer, H., Galil, B. S., Minchin, D., Olenin, S., Amorim, A., Canning-Clode, J., Chainho, P., Copp, G., Gollasch, S., Jelmert, A., Lehtiniemi, M., McKenzie, C., Mikus, J., Miossec, L., Occhipinti-Ambrogi, A., Pecarevic, M., Pederson, J., Quilez- Badia, G., Wijsman, J. & Zenetos, A. (2014) Ten suggestions for advancing assessment and management of nonindigenous species in marine ecosystems. Marine Policy, 44, 160 - 165. https: // doi. org / 10.1016 / j. marpol. 2013.08.019
Poupin, J. (1994) Quelques crustaces decapodes communs de Polynesie Francaise. Rapport scientifique du Service Mixte de Surveillance Radiologique et Biologique de l'homme et de l'environnement, Montlhery Cedex, France, 86 pp.
Sakai, T. (1976) Crabs of Japan and the Adjacent Seas. Vol. 1 - 3. Kodansha Ltd, Tokyo, xxix + 773 pp., 379 figs; 6 + 251 pls.; 461 pp., 2 figs., 3 maps. [Vol. 1: English text: pp. i - xxix + 1 - 773., Figs. 1 - 379; Vol. 2: Pls. 1 - 6 + 1 - 251; Vol. 3: Japanese text: pp. 1 - 461, Figs. 1 - 2, 3 maps]
Stephensen, K. (1945) The Brachyura of the Iranian Gulf with an Appendix: The male pleopod of the Brachyura. Danish Scientific Investigations in Iran. Part 4. E. Munksgaard, Copenhagen, 181 pp. [pp. 57 - 237]

FIGURE 1. Comparison of mitochondrial cytochrome oxidase subunit I (COI) of Gonioinfradens giardi (AP47) and G. paucidentata (AP48). A. Molecular phylogenetic analysis by Maximum Likelihood. B. p-distance table revealing the evolutionary divergence between sequences.

FIGURE 2. Colour in life.Gonioinfradens giardi (Nobili, 1905), male, cl46.5 mm (SMNHTAU-AR 29776), Israel.
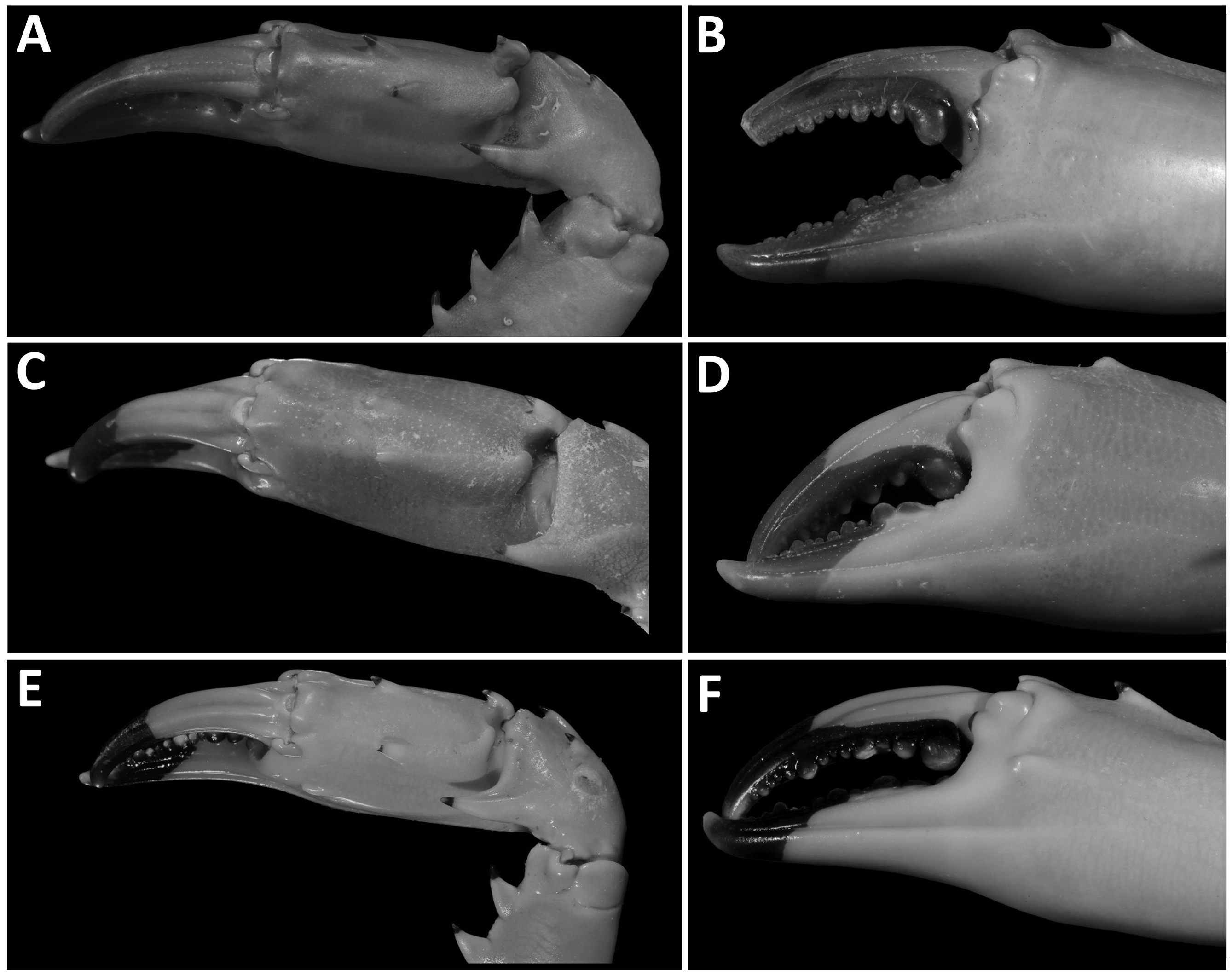
FIGURE 3. Large chela, A, C, E dorsal view; B, D, F, outer view. A, B, Gonioinfradens giardi (Nobili, 1905), male, cl 46.5 mm (SMNHTAU-AR 29776), Israel. C, D, Gonioinfradens giardi (Nobili, 1905), Holotype, male, cl 42.7 mm (MNHN IU- 2014-11969, previouslyMNHN B.5657), United Arab Emirates.E, F, Gonioinfradens paucidentata (A.Milne Edwards, 1861), male, cl 46.9 mm (MNHN), Marquesas Is.

FIGURE 4. Male thoracic sternum and pleon.A, B, Gonioinfradens giardi (Nobili, 1905), male,cl 46.5 mm (SMNHTAU-AR 29776), Israel. C, D, Gonioinfradens giardi (Nobili, 1905), Holotype, male, cl 42.7 mm (MNHN IU-2014-11969, previously MNHN B.5657), United Arab Emirates. E, F, Gonioinfradens paucidentata (A. Milne Edwards, 1861), male, cl 46.9 mm (MNHN), MarquesasIs.

FIGURE 5. Gonopod 1. A, Gonioinfradens giardi (Nobili, 1905), male, cl 46.5 mm (SMNHTAU-AR 29776), Israel. B, Gonioinfradens paucidentata (A. Milne Edwards, 1861), male, cl 46.1 mm (MP B.25967), New Caledonia.

FIGURE 6. Female sternopleonal cavity with vulvae on sternite 6. Gonioinfradens giardi (Nobili, 1905), paralectotype, female, cl 38.3 mm (MNHN IU-2014-11969, previously MNHN B.5657), United Arab Emirates.
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gonioinfradens giardi ( Nobili, 1905 )
| Galil, Bella S., Douek, Jacob, Gevili, Roy, Goren, Menachem, Yudkovsky, Yana, Paz, Guy & Rinekvich, Baruch 2018 |
Gonioinfradens paucidentatus
| Karhan 2012: 118 |
| Corsini-Foka 2010: 331 |
Gonioinfradens paucidentata
| Naderloo 2007: 343 |
| Apel 2001: 72 |
| Apel 1998: 223 |
Charybdis (Gonioinfradens) paucidentata
| Stephensen 1945: 119 |
| Leene 1938: 131 |
Charybdis giardi
| Balss 1924: 3 |
Charybdis (Goniosoma)
| Nobili 1906: 115 |
Thalamita Giardi Nobili, 1905: 164
| Nobili 1905: 164 |
1 (by plazi, 2018-01-15 06:40:59)
2 (by ImsDioSync, 2018-01-15 06:41:41)
3 (by ExternalLinkService, 2019-09-26 01:54:45)
4 (by ExternalLinkService, 2022-01-29 16:14:15)
5 (by ExternalLinkService, 2022-02-06 05:02:11)
6 (by GgImagineBatch, 2022-04-30 04:06:36)
7 (by diego, 2022-11-11 12:42:15)
8 (by ExternalLinkService, 2022-11-12 11:27:34)
9 (by plazi, 2023-10-28 16:07:06)