
Beutelius, Escalona & Lawrence & Ślipiński, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4728.4.11 |
|
publication LSID |
lsid:zoobank.org:pub:B0E44E58-E3AD-40B7-A714-A8C24C0A074D |
|
persistent identifier |
https://treatment.plazi.org/id/A1786336-C244-457F-A8D7-8151BBE86112 |
|
taxon LSID |
lsid:zoobank.org:act:A1786336-C244-457F-A8D7-8151BBE86112 |
|
treatment provided by |
Plazi |
|
scientific name |
Beutelius |
| status |
gen. nov. |
Beutelius gen. nov.
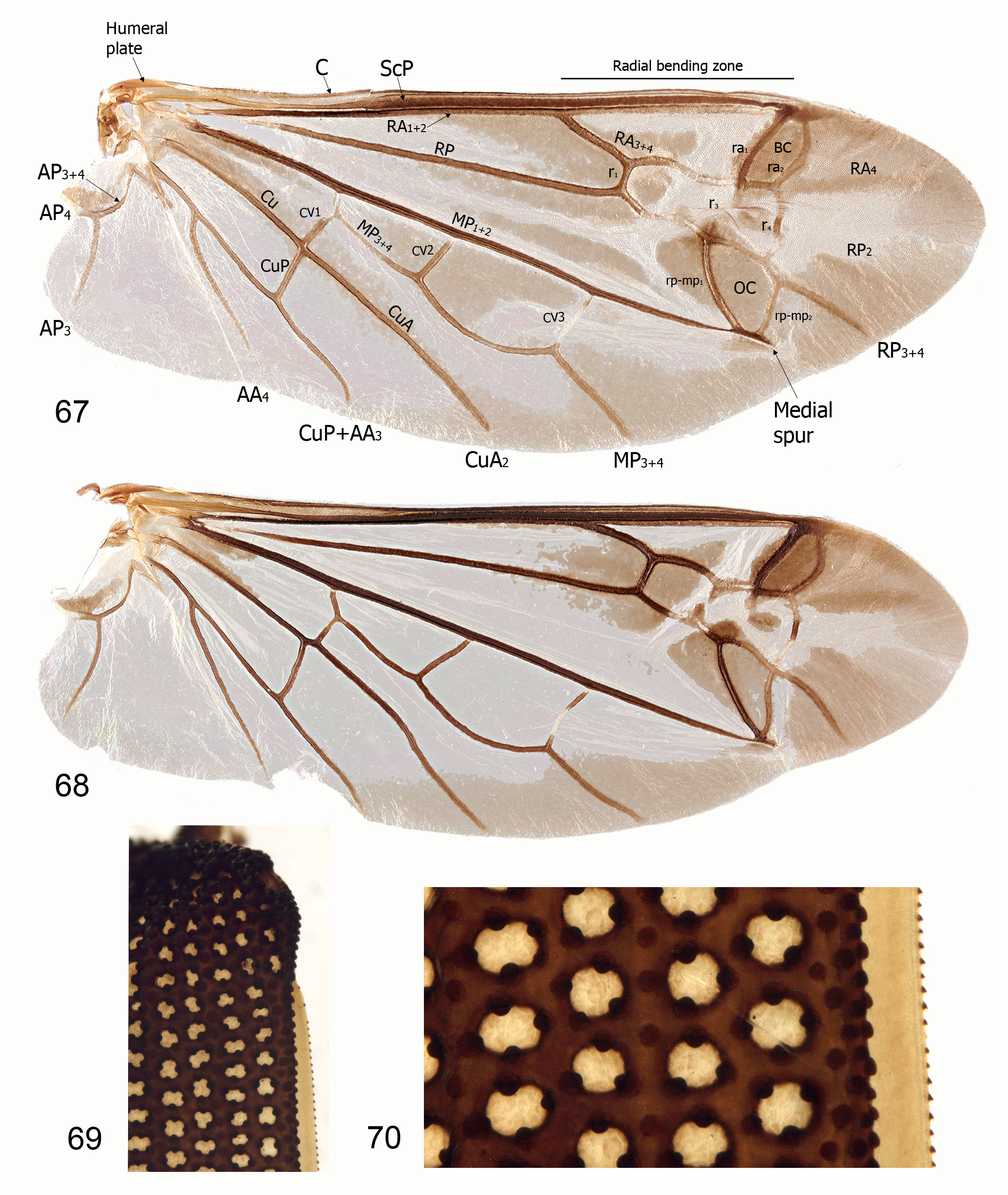
( Figs 4–13 View FIGURES 4–9 View FIGURES 10–13 , 22–38 View FIGURES 22–26 View FIGURES 27–32 View FIGURES 33–38 , 44–50 View FIGURES 39–44 View FIGURES 45–50 , 56–57, 61–62 View FIGURES 56–63 , 67–68 View FIGURES 67–70 , 71–73 View FIGURES 71–75 , 79, 82–89, 91–95 View FIGURES 76–84 View FIGURES 85–95 )
Type species. Omma mastersi MacLeay, 1871 View in CoL .
Diagnosis. Body elongate, moderately flattened, often tuberculate or rarely spinose ( B. rutherfordi ) covered with ribbed scales ( Figs 4–13 View FIGURES 4–9 View FIGURES 10–13 ). Head longer than wide, narrowed behind eyes and with distinct neck region ( Figs 22 View FIGURES 22–26 , 27 View FIGURES 27–32 , 33 View FIGURES 33–38 ). Ligula rounded, labial and maxillary palps elongated, when extended posteriorly clearly reaching beyond eye level. Posterior tentorial pits not evident. Anterior third of gulamentum depressed to accommodate labial and maxillary palps ( Figs 23 View FIGURES 22–26 , 28 View FIGURES 27–32 ), very slightly in B. rutherfordi ( Fig. 34 View FIGURES 33–38 ). Antennae filiform to moniliform ( Figs 37, 38 View FIGURES 33–38 ), rarely extending beyond base of prothorax ( B. rutherfordi ) and setose. Prothorax without lateral pronotal carinae; notopleural suture complete to anterior edge, sternopleural suture present ( Fig. 49 View FIGURES 45–50 ) or absent ( B. rutherfordi , Fig. 44 View FIGURES 39–44 ). Prosternal process extending between procoxae ( Fig. 48 View FIGURES 45–50 ). Epipleura narrow ( B. rutherfordi ) to broad and complete; elytral sutural flange finely denticulate as part of the interlocking mechanism (fig. 11 in Lawrence 1999). Outer surface of protibia in male with ( Figs 79, 81, 82 View FIGURES 76–84 ) or without ( Fig. 84 View FIGURES 76–84 ) setose excavation at basal third.
Description. Length: 6–26 mm. Body 2.4–3.0 times as long as wide, slightly flattened dorsally and distinctly so ventrally, with sides of pronotum and elytra slightly, independently rounded or pointed ( B. rutherfordi ). Colour uniformly dark brown to black; upper surfaces tuberculate, tubercles spine-like in B. rutherfordi , and clothed with flattened, ribbed scales which differ in colour and form a distinct pattern (interspersed with stout, erect spines in B. rutherfordi ); last abdominal ventrite in males often with longer and denser setae.
Head prognathous, constricted posteriorly to form a neck, temples often present (indistinct in B. rutherfordi ), median occipital endocarina present. Eyes entire and very finely facetted, without interfacetal setae. Antennal insertions lateral, barely concealed by frontal ridges; subantennal groove absent. Frontoclypeal suture absent. Labrum transverse or slightly transverse, apically convex, fused to clypeus. Antenna usually short, rarely extending beyond base of prothorax ( B. rutherfordi ), 11-segmented, filiform to moniliform. Mandible tridentate with vertically aligned teeth, without mola or prostheca; its dorsal surface near base with setose cavity. Maxilla highly reduced and partly concealed by mentum ( Fig. 20 View FIGURES 14–21 ); cardo and stipes sclerotised; galea and lacinia slender, hyaline and setose, the latter without uncus. Maxillary palpomeres 1, 2 and 3 with a pair of long apical setae, apical palpomere strongly expanded and securiform with small cavity bearing sensilla (figs 5, 7 in Lawrence 1999) near outer edge of upper surface. Mentum longer than wide or about as long as wide ( B. rutherfordi ) with deep basal pit, barely ( B. mastersi ) to clearly ( B. sagitta ) concave laterally; ligula small and rounded, located on dorsal surface and concealed by mentum together with palpal insertions in ventral view. Labial palpomeres 1 and 2 with a pair of long apical setae, apical palpomere strongly expanded and securiform, with a cavity like that on the apical maxillary palpomere. Posterior tentorial pits ventral, reduced and not clearly visible. Gular sutures well separated; gula longer than wide. Anterior third of gulamentum depressed to accommodate labial and maxillary palps, weakly depressed in B. rutherfordi . Corpotentorium weakly developed or absent. Cervical sclerites absent.
Prothorax usually distinctly narrower than the distance across elytral humeri (less so in the male of B. rutherfordi ), widest anteriorly with sides slightly sinuate; lateral pronotal carinae absent (irregular in B. rutherfordi ). Pronotal disc slightly convex or concave ( B. sagitta ) with paired anterior and posterior deep impressions; the anterior impressions closer together and the posterior ones closer to the posterior angles. Prosternum in front of coxae moderately long, partly or completely fused to pleuron (pleurosternal suture absent in B. rutherfordi ), without paired crural impressions. Prosternal process incomplete, extending between procoxae, narrowed apically and acute at apex. Notopleural suture complete to anterior edge, never joined by pleurosternal suture. Procoxae subglobular, not strongly projecting, without concealed lateral extensions, without or with reduced coxal plates; trochantin large, well sclerotised and broadly exposed. Procoxal cavities slightly transverse, contiguous, broadly open externally and internally. Promesothoracic interlocking mechanism formed by mesanepisternal condyle and propleural cavity. Scutellar shield distinctly elevated above the anterior portion of the mesoscutellum, not elevated above elytra, sides parallel ( Fig. 87 View FIGURES 85–95 ). Elytra complete with independently rounded to acute apices and ten rows of circular window punctures; elytral sutural edge with flange margin finely denticulate as part of the interlocking mechanism, the denticles sparser in B. rutherfordi (fig. 11 in Lawrence 1999); epipleura very narrow to broad and complete. Mesoventrite at middle with acute anterior projection separating paired procoxal rests and extending onto mesanepisterna, mesoventral cavity absent; discrimen short and transverse (mesokatepisternal) suture incomplete (not always visible externally). Mesoventral process divided into two short, acute processes. Mesocoxae more or less globular, slightly projecting, without or with short plates and broadly exposed trochantins. Mesocoxal cavities slightly transverse, contiguous, partly closed laterally by mesepimera and metanepisterna; meso- and metathoracic joint within coxal cavities membranous. Metaventrite narrowed anteriorly; discrimen present; transverse (metakatepisternal) suture well developed. Metanepisternum completely exposed, widest near anterior end; exposed portion of metepimeron narrowly elongate and widest at posterior end; metatrochantins exposed. Metacoxae strongly transverse with subconical mesal projections, contiguous, extending laterally to meet elytra, with weak coxal plates. Metendosternite with a moderately long, slender stalk, short, broad ventrolateral processes, moderately long and distally slender lateral arms, moderately long, and an apically emarginate anterior process with tendons moderately widely separated on either side of emargination.
Hind wing often well-developed and moderately broad (reduced in males of B. rutherfordi ), with very short bending zone near apex of radial bar (anterior wing strut); pterostigma absent; two cross-veins join RA 1+2 and RA 3+4 to form brachial cell ( Atkins 1963); RP complete almost to wing base, joined to RA 3+4 by three cross-veins; RP and MP joined by two cross-veins forming the oblongum cell; MP 1+2 with very short bending zone or hinge and a very short, straight medial spur; medial field with four free veins and no medial fleck; wedge cell absent; anal lobe well developed, without embayment; AP 3+4 forked.
Legs short and slender; tibia subequal in length to femur; protibia posterior side with setose excavation ( B. rutherfordi ) or pit ( B. sagitta ) at basal third in males, in B. mastersi female protibia basal third with a patch of black ribbed scales ( Fig. 6 View FIGURES 4–9 ); protibial apex not or slightly expanded ( B. mastersi ). Tibial spurs well-developed, usually unequal and paired on all legs; protibial spurs scoop-like and widely separated, major mesotibial spur scoop-like, metatibial spurs pointed to apex and subequal. Tarsi 5-5-5; tarsomeres without ventral lobes; pretarsal claws simple; empodium bisetose.
Abdomen ( Figs 71, 72, 73 View FIGURES 71–75 ) with five flattened ventrites more or less connate, separated by narrow grooves and all on the same plane; lateral edges of ventrites 1 to 4 with small bare patches, ventrite 5 in males with longer and fine, yellow hairs, posterior edge often not margined. Aedeagus of modified trilobate type, with parameres completely fused to phallobase and partially fused together, each with a mesal notch near apex. Ovipositor narrowly elongate, gonocoxites each with well-developed apical gonostylus.
Etymology. The genus is named after our dear friend Rolf Beutel (Jena, Germany) to commemorate his remarkable career in insect systematics. Gender: masculine.
Species. B. mastersi (MacLeay) , comb. nov., B. rutherfordi (Lawrence) , comb. nov., B. sagitta (Neboiss) , comb. nov. and B. reidi sp. nov.
Comments. Lawrence (1999) noted that Omma was heterogeneous morphologically and if the genus were to be split it would be logical to remove the scale-covered species from Omma . This division was also recovered as two distinct clades in a morphological phylogenetic analysis by Hörnschemeyer (2009); see comments under Omma description. Only recently we were able to study additional specimens of each species, including the male and female of B. sagitta and B. mastersi respectively. This new material and a detailed morphological examination allowed us to identify new diagnostic characters, such as the anterior depression in the gulamentum ( Figs 23 View FIGURES 22–26 , 28 View FIGURES 27–32 ), that separate Beutelius from Omma without ambiguity. Unfortunately, this critical character is not easily observable in compression fossils and the Omma fossil species could not be placed with certainty. A careful study of these extinct taxa is necessary to classify them based on reliable diagnostic characters.
Biology. A presumed Beutelius larva, based on a single specimen, was described by Lawrence (1999) and Grebennikov (2004) from Red Hill in southern WA. The species could belong to B. sagitta or B. rutherfordi . Adult B. mastersi have been observed active on bark during the day, their colouration resembles that of other local insects (e.g., the beetle Stigmatium gilberti White and the mutillid wasp Ephutomorpha sp.) and they may be part of a mimicry complex ( Lawrence 1999). The distinctive male setose excavation in the basal third of the protibia is absent in B. mastersi . Curiously, some females of this species ( Figs 6, 7 View FIGURES 4–9 , 26 View FIGURES 22–26 , 31 View FIGURES 27–32 , SAMA-25-038603) have black, ribbed, scale-like setae in the same area, but the biological implications of this feature are unknown. Beutelius rutherfordi females are fully winged while males are micropterous, a rare feature in beetles ( Lawrence 1999).
Distribution, Beutelius mastersi is known from inland NSW and QLD. Beutelius rutherfordi and B. sagitta are known from the Mallee woodlands and shrub lands of SA and WA. Beutelius reidi is only known from the type locality in northwest New South Wales.












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.