
Pulvinaria psidii Maskell, 1893
|
publication ID |
https://doi.org/10.11646/zootaxa.4868.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:AD0B95D7-70AF-4B07-A7A7-F02F5DC4934D |
|
DOI |
https://doi.org/10.5281/zenodo.4417892 |
|
persistent identifier |
https://treatment.plazi.org/id/DD2B87BF-FF9F-FFE7-06D6-25EE74DECE83 |
|
treatment provided by |
Plazi |
|
scientific name |
Pulvinaria psidii Maskell, 1893 |
| status |
|
Pulvinaria psidii Maskell, 1893 View in CoL
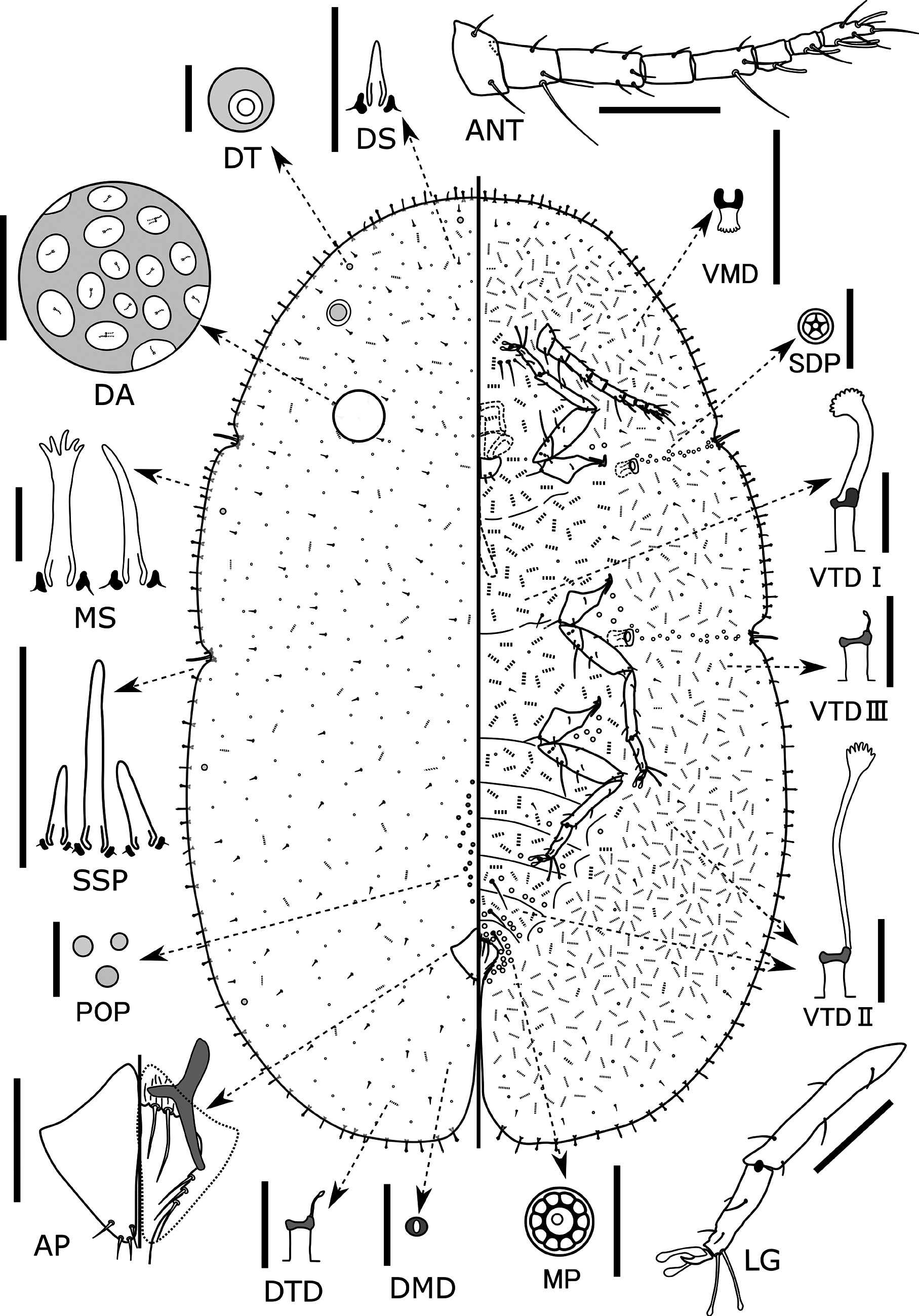
( Fig.3 View FIGURE 3 )
Pulvinaria psidii Maskell, 1893: 223 View in CoL ; Green, 1909: 258, 264; 1928: 24; Fernald, 1903: 137; Steinweden, 1946: 11; Zimmerman, 1948: 333, 336; Beardsley, 1966: 492; Hodgson, 1968: 168; Ezzat & Hussein, 1969: 228; Kawai, 1980: 152; Tao et al., 1983: 85; Hamon & Williams, 1984: 102; Williams & Williams, 1988: 59; Williams & Watson, 1990: 153; Qin & Gullan, 1992: 144; Ben-Dov 1993: 278; Kawai, 2003: 305, 511, 558.
Material examined. JAPAN: Okinawa Prefecture, Okinawa Is., Naha-shi, Syurisakiyama-cho , on Bischofia javanica , 13. xii.1999, coll. T. Uesato, 4 adult females mounted singly (2 EUMJ, 2 ELKU) .
Redescription. Live appearance: adult female elongate oval, usually flat. Dorsum dark yellow to greenish yellow, dorsum without visible wax before oviposition, but developing a small amount of granular or filamentous white wax during the oviposition period. Ovisac short, about 1–2 times as long as body; posterior part of body strongly lifted by ovisac.
Slide-mounted adult female (n=4): body elongate oval, 2.2–3.0 mm long, 1.3–1.9 mm wide, margin with a distinct and relatively deep indentation at each stigmatic cleft; anal cleft approximately 1/4–1/6 of body length.
Dorsum. Derm membranous and dermal areolations well developed. Dorsal setae spiniform, frequent, scattered over entire dorsum, each 3–6 µm long with a well-developed basal socket. Preopercular pores oval to circular, each 2–3 µm in diameter, barely sclerotised and often difficult to see and count, with 17–24 present anterior to anal plates. Tubular ducts and microducts frequent throughout, each one situated within an areolation ( Fig. 3 View FIGURE 3 DA). Dorsal tubercles of normal convex type present in submarginal areas, with 2–4 between anterior stigmatic clefts, each side with 0 or 1 between anterior and posterior stigmatic clefts, and 1–3 between posterior stigmatic cleft and anal cleft. Anal plates together quadrate; each plate 140–160 µm long, 70–82 µm wide, with a well-developed and bifurcate supporting bar, a slightly convex posterolateral margin and 3 or 4 apical setae. Ano-genital fold with 2 pairs of setae along anterior margin and 2 or 3 pairs laterally. Anal ring bearing 6 setae. Eyespots situated in submarginal area.
Margin. Marginal setae with well-developed basal sockets, mostly with strongly fimbriate apices; length of each seta 10–28 µm, each side with 14–19 setae between anterior and posterior stigmatic clefts. Stigmatic clefts distinct and rather deep, each containing 3 stigmatic spines, median spine longest, 30–60 µm long, approximately 2–4 times as long as a lateral spine.
Venter. Derm membranous. Multilocular pores each 5–8 µm wide, with 9–12 (usually 10) loculi, present around genital opening and on medial areas of preceding 4 or 5 abdominal segments; a small group also present lateral to each coxa. Spiracular pores each 3–4 µm wide, mostly each with 5 loculi, present in rather narrow bands 1–3 pores wide between margin and each spiracle; anterior bands each containing 13–24 pores, posterior bands each with 17–34 pores. Microducts scattered throughout venter. Tubular ducts of three types: type I each with large outer ductule ( 2–3 µm wide and 5–7 µm long), a stout inner ductule ( 2–3 µm wide and 15–20 µm long) and a well-developed flower-shaped terminal gland, present in posterior-medial area of head, medial areas of all thoracic segments and anterior abdominal segments, also in inner submarginal area of head, thorax and anterior abdominal segments; type II tubular ducts each with large outer ductule ( 2–3 µm wide and 5–10 µm long) and a shallow cup-shaped invagination leading to an extremely long and narrow inner ductule (< 1 µm wide and 25–45 µm long) with a well-developed terminal gland, mostly occurring in medial area of the posterior abdominal segments and submarginal area of head, thorax and abdomen; type III ducts similar to type II but each with a short, filamentous inner ductule (< 1 µm wide and 1–3 µm long) and minute terminal gland, present in a broad submarginal band on head, thorax and abdomen and forming a complete submarginal ring, intermixed with type I ducts in inner submarginal areas of head, thorax and anterior abdominal segments and with type II ducts in broad outer submarginal area on head, thorax and abdomen. Posterior 3 abdominal segments each with 1 pair of long ventral setae in medial area. With 4 or 5 pairs of long setae present between antennal bases and 0–2 pairs of long setae present on area mesad of each coxa; other setae short and fine, distributed over entire venter. Spiracles normal for the genus but, in mature specimens, each surrounded by a well-developed sclerotised spiracular plate; peritreme widths: anterior spiracle 30–42 µm, posterior spiracle 40–54 µm. Legs well developed, each with a completely articulated tibio-tarsal joint and an articulatory sclerosis; claw without a denticle; both claw digitules rather broad and slightly shorter than thin tarsal digitules. Hind trochanter + femur 240–294 µm long, hind tibia 181–230 µm long, and hind tarsus 103–122 µm long. Antennae each 6 to 8segmented (mostly 8), 319–432 µm long. Labium 85–110 µm wide, 52–72 µm long.
Host-plants in Japan. Anacardiaceae : Mangifera indica ( Kawai 2003) ; Apocynaceae : Plumeria rubra ( Kawai 1980) ; Moraceae : Ficus boninsimae ( Kawai 1980) ; Myrsinaceae : Ardisia sieboldii ( Kawai 1980) ; Myrtaceae : Psidium guajava ( Kawai 1980, 2003), Syzygium buxifolium ( Kawai 1980) , S. cleyerifolium ( Kawai 1980) ; Phyllanthaceae : Bischofia javanica ( Kawai 1980) ; Pittosporaceae : Pittosporum boninense ( Kawai 1980) ; Rubiaceae : Morinda citrifolia ( Kawai 1980) , Tarenna subsessilis ( Kawai 1980) ; Rutaceae : Citrus spp. ( Kawai 2003); Sapotaceae : Planchonella obovata ( Kawai 1980) ; Theaceae : Schima wallichii ( Kawai 1980) ; Ulmaceae : Trema orientalis ( Kawai 1980) ; Verbenaceae : Callicarpa glabra ( Kawai 1980) . Worldwide host-plants records of this species are given by García Morales et al. (2016).
Remarks. Pulvinaria psidii is similar to P. aurantii and P. polygonata in having fimbriate marginal setae, welldeveloped dermal areolations and eyespots situated in the submarginal area of the dorsum. However, it differs from them in having (contrasting character states in P. aurantii and P. polygonata in parentheses): (i) multilocular pores mostly each with 10 loculi (mostly each with 7 loculi), (ii) an extremely long inner ductule (about 5–10 times as long as outer ductule) in type II ventral tubular ducts (inner ductule of type II ventral tubular ducts mostly 2–3 times as long as outer ductule), and (iii) in having a bifurcate supporting bar on underside of each anal plate (supporting bars not bifurcate). Important diagnostic morphological character states of P. psidii plus a comparison with the type species of the genus, P. vitis , are summarised in Table 1. Pulvinaria psidii can be separated from other Pulvinaria species distributed in Ryukyu Islands and P. vitis by condition of dermal areolation, location of eyespots, shape of marginal seta apices, Type III ventral tubular duct distribution, the number of loculi in each multilocular pore, and the number of stigmatic spines in each stigmatic cleft.
The adult female morphology of P. psidii described here mostly agrees with the redescription of the species by Qin & Gullan (1992) but with slight differences, as follows (character states in Qin & Gullan in parentheses): (i) preopercular pores numbering 17–24 (22–49); and (ii) anal ring setae numbering 6 (8). These morphological discrepancies may be due to regional morphological variation in the species.
| EUMJ |
Ehime University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pulvinaria psidii Maskell, 1893
| Tanaka, Hirotaka & Kamitani, Satoshi 2020 |
Pulvinaria psidii
| Kawai, S. 2003: 305 |
| Ben-Dov, Y. 1993: 278 |
| Qin, T. K. & Gullan, P. J. 1992: 144 |
| Williams, D. J. & Watson, G. W. 1990: 153 |
| Williams, J. R. & Williams, D. J. 1988: 59 |
| Hamon, A. B. & Williams, M. L. 1984: 102 |
| Tao, C. C. C. & Wong, C. Y. & Chang, Y. C. 1983: 85 |
| Kawai, S. 1980: 152 |
| Ezzat, Y. M. & Hussein, N. A. 1969: 228 |
| Hodgson, C. J. 1968: 168 |
| Beardsley, J. W. 1966: 492 |
| Zimmerman, E. C. 1948: 333 |
| Steinweden, J. B. 1946: 11 |
| Green, E. E. 1909: 258 |
| Fernald, M. E. 1903: 137 |
| Maskell, W. M. 1893: 223 |