
Ischnocnema lactea ( Miranda-Ribeiro 1923 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4444.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:4CCE3CF1-A111-40FB-9518-5D6EAD02A5C0 |
|
DOI |
https://doi.org/10.5281/zenodo.5960219 |
|
persistent identifier |
https://treatment.plazi.org/id/DE3D87CC-FFD7-FFB7-F8C7-FE30FC3BFF21 |
|
treatment provided by |
Plazi |
|
scientific name |
Ischnocnema lactea ( Miranda-Ribeiro 1923 ) |
| status |
|
Ischnocnema lactea ( Miranda-Ribeiro 1923) View in CoL
Basanitia lactea Miranda-Ribeiro 1923: 851 View in CoL
Eleutherodactylus lacteus —Lynch 1968: 875
Eleutherodactylus (Eleutherodactylus) lacteus — Lynch & Duellman 1997: 226
Ischnocnema lactea View in CoL —Heinicke et al. 2007: 15; Hedges et al. 2008: 27
Lectotype. MZUSP 828; adult female, collected by Gustavo Edwall from the Municipality of Iguape , State of São Paulo, Brazil. The original description did not provide a date or specific sampling locality (see discussion for an analysis on the type locality). Bokermann (1966) designated the lectotype .
Paralectotype. MZUSP 504; adult male of Ischnocnema parva (Girard 1853) from Campo Grande, Municipality of Santo André, State of São Paulo, Brazil. The paralectoype was designated by Bokermann (1966) and later redetermined by Heyer (1985) as Ischnocnema parva .
Type locality. Municipality of Iguape , State of São Paulo, Brazil. Restricted by Bokermann (1966).
We note, however, that the record for the Municipality of Iguape, which was formerly considered the type locality, is in error. Here, we redefine the type locality as the area surrounding the Padre Dória road, in the Juqueriquerê River basin (coordinates 23°38'32.43"S, 45°41'39.20"W, datum = WGS84; altitude 1,010 m a.s.l.), Municipality of Caraguatatuba, State of São Paulo, Brazil (see discussion for details).
Examined specimens. A total of 47 specimens of I. lactea were examined, which included 18 adult males, 13 adult females (including the lectotype; Fig. 2 View FIGURE 2 ), 13 sub-adults, two young individuals, and one adult of undetermined sex. Specimens were obtained from different populations inhabiting the Serra do Mar region in the State of São Paulo, Southeastern Brazil: Núcleo Curucutu , PESM, Municipality of Itanhaém (N= 7), and Municipality of São Paulo (N= 1); Núcleo Santa Virgínia , PESM, Municipality of São Luís do Paraitinga (N= 1); Serra da Cantareira, Municipality of São Paulo (N= 1); Reserva Florestal do Morro Grande, Municipality of Cotia (N= 2); Estação Biológica de Boracéia (EBB), Municipality of Salesópolis (N= 2) ; PNMNP, Municipality of Santo André (N= 21); Parque Estadual de Ilhabela ( PEI), Municipality of Ilhabela (N= 2), and Municipality of Iguape (type locality; N= 1, lectotype). Juveniles ( MZUSP 134037 View Materials , 145253 View Materials ) and sub-adults (four males, CFBH 11171 View Materials , 16772 View Materials , 29118 View Materials , ZUEC 6382 View Materials ; nine females, CFBH 17670 View Materials , 29094 View Materials , 29107-08 View Materials , MZUSP 145260 View Materials , ZUEC 3531 View Materials , 6381 View Materials , 6383 View Materials , 6470 View Materials ) were not considered in morphological and morphometric comparisons. Moreover , lectotype MZUSP 878 was in moderate condition of preservation ( Fig. 2 View FIGURE 2 ).
Diagnosis. Ischnocnema lactea is diagnosed by the following combination of characters: (1) large-sized species (among I. lactea species), with adult males SVL 19.6–26.7 mm; X = 23.3. ± 2.4 mm; N = 18; adult females SVL 27.6–34.3 mm; X = 30.2 ± 2.5 mm; N = 10; (2) dorsum, lateral, and gular region rugose with several scattered tubercles; (3) venter moderately granular; (4) head longer than wide; (5) snout rounded in dorsal view; (6) tip of snout obtuse in lateral view; (7) eyelid tubercles oval, well-developed; (8) vomerine process present, bearing prevomerine odontoids; (9) vocal sac single, subgular, externally expanded; (10) ungual flap present, disks expanded and indented, T-shaped, truncate in fingers II–IV; (11) double glandular nuptial pad; (12) toes relative length I<II<III<V<IV; (13) calcar tubercle present, moderate; (14) in life, iris varying from golden to lemonyellowish, with a medial reddish band crossing the black pupil; (15) in life, hidden areas of inguinal region, thighs, shanks, and tarsus show a large array of patterns and bright colors; (16) aforementioned bright colors may appear on marbled pattern in a dark background; (17) black blotch on cloacal region, tarsus, and feet.
Comparison with other species of Ischnocnema lactea . Ischnocnema lactea is distinguishable from all other species of I. lactea as follows (character states for other members in parentheses): large-sized species with adult males reaching SVL 19.6–26.7 mm, and adult females SVL 27.6–34.3 mm (adult males are smaller in I. concolor 14.7–18.4 mm, I. gehrti 14.2–15.3 mm, I. melanopygia 15.7–18.7 mm, I. paranaensis 17.7 mm, I. randorum 11.8– 15.0 mm, I. spanios 14.7 mm, and I. vizottoi 13.3–17.0 mm; overlap in size on adult males of I. holti 18.4–24.9 mm and I. nigriventris 18.2–20.2 mm); dorsum very rugose, with several tubercles and granules (dorsum smooth in I. concolor , I. gehrti , I. melanopygia , I. paranaensis , I. randorum , and I. spanios ); snout rounded in
dorsal profile (subovoid in I. gehrti , I. melanopygia , I. randorum , I. spanios , and I. vizottoi ; rounded in the other species); tip of snout may present a medial process, providing a subovoid aspect in dorsal view (variation also occurs in I. concolor and I. vizottoi ); tip of snout obtuse in lateral view (also obtuse in I. holti ; truncate in I. gehrti and I. spanios ; rounded in other species); eyelid tubercles oval well developed (absent in I. concolor ; oval poorly developed in I. gehrti , I. holti , I. melanopygia , I. randorum , and I. spanios ; conical well developed in I. nigriventris ; unknown in I. paranaensis ); patches of prevomerine odontoids present (absent in I. paranaensis and I. randorum ); vocal sac single, subgular and externally expanded (not expanded in I. gehrti , I. holti , I. spanios , and I. melanopygia ; unknown in I. paranaensis ); in life, iris varies from golden to lemon-yellowish, with a medial stripe varying from reddish to orange, crossing the black pupil (golden to lemon-yellowish, with brown to black medial stripe in I. concolor , I. holti , I. melanopygia , I. randorum , and I. vizottoi ; iris silver with a ventral yellowish copper band in I. nigriventris ; dorsally bluish in I. spanios ; not reported for I. gehrti and I. paranaensis ); disks expanded, T-shaped, truncate in fingers II–IV (similar for most species; disks expanded Tshaped, truncate in fingers III–IV in I. concolor , I. melanopygia , and I. vizottoi ; disk finger II rounded in I. melanopygia , I. vizottoi and, I. concolor , slightly expanded in the latter); double glandular nuptial pads present (single glandular in I. concolor , I. gehrti , I. melanopygia , I. randorum , I. spanios , and I. vizottoi ; unknown in I. paranaensis ); toes relative length I<II<III<V<IV (similar in most of species; I<IIẼV<III<IV in I. gehrti , I. melanopygia and I. spanios ); calcar tubercles moderate (also moderate in I. randorum , I. spanios , and I. gehrti ; poorly developed in I. holti , I. concolor , and I. vizottoi , occasionally inconspicuous in the last two species; well developed in I. melanopygia and I. nigriventris ; absent in I. paranaensis ); in life, inguinal region and hidden areas of thigh, shank, and tarsus show a large array of bright colors and patterns (bright coloration on mentioned areas absent in I. concolor , I. paranaensis , and I. vizottoi ); bright colors on mentioned areas appear on marbled pattern in a dark background (pattern does not occur in I. concolor , I. gehrti , I. paranaensis , I. randorum , and I. spanios ); black blotch on cloacal region, tarsus, and feet present (absent in most of the species, except in I. gehrti , I. melanopygia , and I. spanios ; unknown for I. paranaensis ).
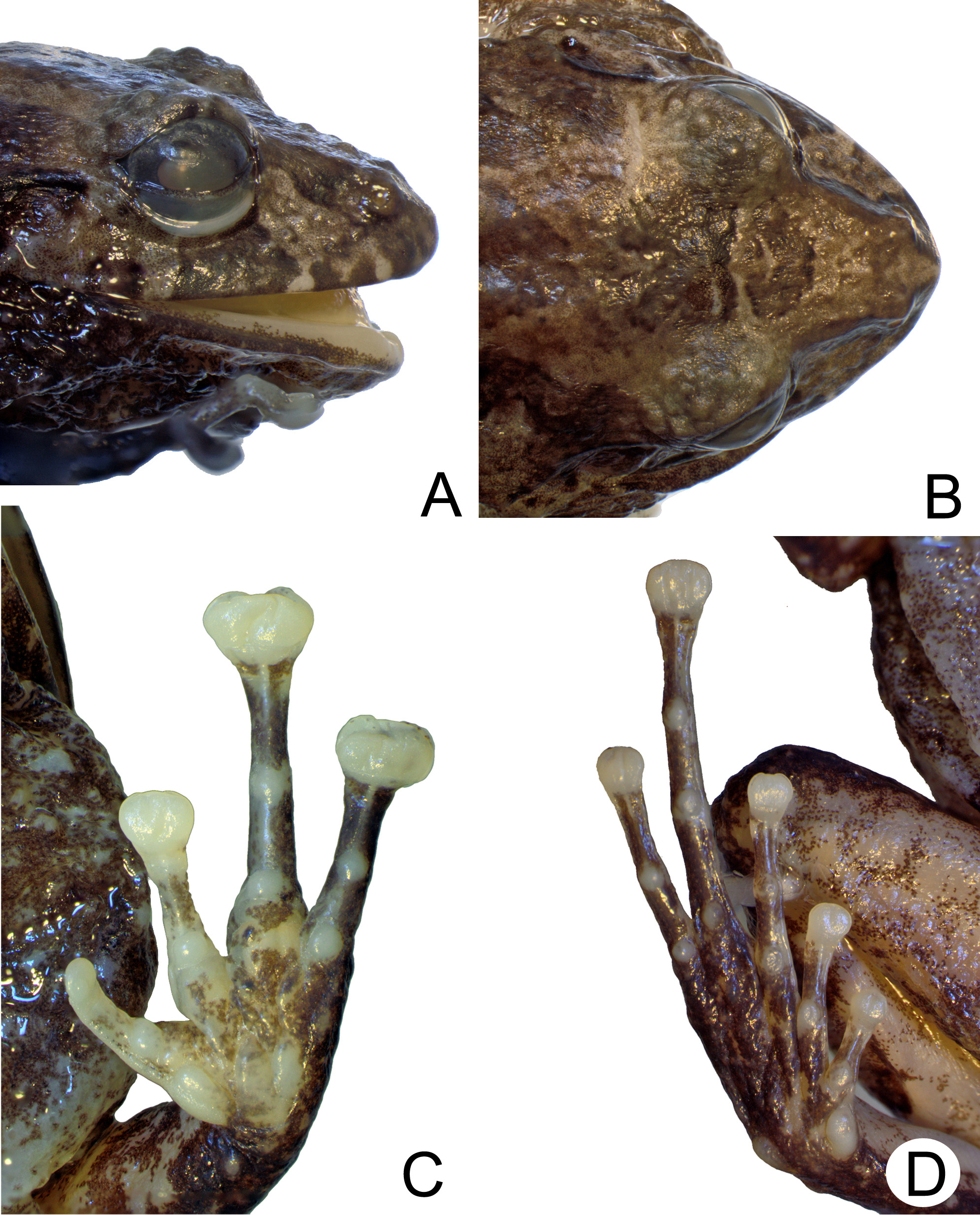
Redescription of the lectotype MZUSP 828 View Materials . According to original description ( Miranda-Ribeiro 1923), adult female from the Municipality of Iguape , State of São Paulo, Southeastern Brazil, collected by Gustavo Edwall . The specimen shows a moderate condition of preservation, having lost some general characters. The lectotype has a robust body, dorsum smooth, presenting very few small, scattered tubercles; flanks and ventral surfaces granular (bely and laterals) to very granular (cloacal region); skin translucent, with muscles seen by transparency mainly on arms and legs. Venter almost entirely open with organs exposed; pectoral and abdominal folds indistinct. Head slightly longer than wide, head length 37% of SVL; snout acuminate in lateral view and subovoid in dorsal; upper jaw show a large medial cut, and projects beyond lower jaw; large eyes, anterolaterally oriented; pupil horizontal and elliptical; translucent palpebral membrane, faintly pigmented on edge; both eyelids present some tuberculation mainly on edges; tympanum moderately distinct, with membrane hidden beneath skin and annulus evident only ventrally; supratympanic fold poorly developed, probably shabby; three medium to large-sized postcommissural tubercles on both side of head. Canthus rostralis distinct, straight; loreal region oblique, dorsolaterally directed and slightly concave; nostrils oval, protruding in dorsal and lateral views, posterolaterally directed; inter-nostril distance 22.7% of HW; choanae nearly rounded, distant from one; prevomerine processes with odontoids present, oblique, situated between and posterior to the choanae; thick, medium-sized tongue, elliptically disposed on mouth floor, notched medially. Arms and forearms moderately slender. Fingers long, with all finger tips lost, which does not allow comparisons with the original state; subarticular tubercles single, oval, well developed; outer metacarpal tubercle cordiform and inner metacarpal tubercle oval, both worn out; supernumerary tubercles on palmar region absent or inconspicuous. Legs moderately slender; thigh almost as long as shank; feet with worn conical outer metatarsal tubercle, and elliptical inner metatarsal tubercle with detached external edge; both inner and external metatarsal tubercles moderately developed; subarticular oval tubercles moderately developed; supernumerary tubercles absent or inconspicuous. Toes long, with some toes tips lost, not allowing to state their original relative length; toes not webbed nor fringed, with ungual flaps present; toes I–V with disk expanded, rounded, dorsally emarginated, indented; disk pads even, broadened, following their disks sizes; calcar inconspicuous.
Measurements of lectotype MZUSP 828 (in mm). SVL 30.5; HL 11.4; HW 11.3; ED 4.4; END 2.8; ESD 4.9; IOD 2.8; IND 2.6; TD 1.3; FAL 6.5; HAL 7.5; THL 13.8; SHL 15.3; FL 19.0, and 4TL 7.8, both considering the lost of 4TD.
Color in preservative. General color of body varying from cream to milky. Few blotches, dots, and stripes can still be observed along the body surfaces. Some light brown blotches are present on dorsum, laterals, head, and dorsal region of thighs.
Intraspecific variation based on newly collected adult specimens. In general, specimens show a body robust, dorsum very rugose, presenting several tubercles of varied sizes. Flanks and undersurfaces varied from granular (venter, arms, and legs) to very granular (lateral, cloacal, and gular regions). In comparison to live individuals, fixed specimens might present body surfaces much less rugose and granular, probably due to fixation. Rugosity also varies in intensity on different parts of the body of the same specimen. Pectoral fold tenuous, lined from axilla to axilla, positioned slightly below the arm–axilla joint; abdominal fold distinct, more conspicuous at the posterior portion of belly than laterals. Head longer than wide, snout acuminate in lateral view, and subovoid in dorsal view mainly due to a medium to large tubercle that can be present at the tip of snout (e.g., CFBH 29055; MZUSP 145317); upper jaw projecting beyond lower jaw; large eyes, protuberant when opened, anterolaterally oriented, with pupil horizontal and elliptical; translucent palpebral membrane, pigmented on edge; eyelids vary from presenting none to several oval tubercles well developed; tympanum moderately distinct, with membrane hidden beneath skin and annulus evident only ventrally; supratympanic fold moderately developed; three to four medium to large-sized postcommissural conical tubercles can be present on both side of head.
There are some small to medium-sized conical tubercles that form the end of supratympanic fold. Canthus rostralis distinct, straight; loreal region oblique, dorsolaterally directed and slightly concave; nostrils oval, protruding in dorsal and lateral views, posterolaterally directed; inter-nostril distance average 27.1% of HW; choanae nearly rounded, distant from one another; prevomerine processes present, oblique, well developed (generally twice its width), situated between and posterior to the choanae, with about four to six odontoids on each patch; premaxillary teeth present; thick, medium-sized tongue, elliptically disposed, notched medially; vocal slits large, conspicuous, anterolaterally oriented, beginning anteriorly to the tongue notch, and ending anteriorly to the line of buccal commissure. Vocal sac single, largely expanded, covering all over the gular region, dorsally visible, however, some males may show vocal sacs not expanded (e.g., CFBH 1936, 28167) and, in general, the gular region of males is darker than in females. Some adult males with a dark gular region may also show a lighter medial line (e.g., CFBH 19368, 29055, 29082).
Arms and forearms moderately robust; three to five lined-up tubercles moderately developed are present on ventral surface of each forearm. Fingers long, not fringed, with ungual flaps present; relative length of fingers I<II<IV<III; finger III average length 68.5% of hand length; subarticular tubercles generally single, oval, well developed; outer metacarpal tubercle cordiform, well developed; inner metacarpal tubercle oval, well developed; supernumerary tubercles on palmar region oval, poorly developed, some of them grayish; double glandular nuptial pad present, beginning on the middle-dorsal surface of finger I, extending posteriorly to the inner metacarpal tubercle, covering half of their surface. Disk largely expanded and T-shaped, truncate on fingers II–IV, with dorsal surface emarginated and medially indented; disk pads even and broadened. Disk on finger I neither expanded nor emarginated, being rounded with indentation absent or indistinct; disk pads even, ovoid, following their disks sizes.
Legs moderately robust; thigh almost as long as shank, which is slightly longer. Dorsolateral surfaces of thighs, shanks, tarsus, and feet with some poorly-developed, scattered tubercles; feet with conical outer metatarsal tubercle and elliptical inner metatarsal tubercle with thick detached external edge; both inner and external metatarsal tubercles moderately developed; subarticular oval tubercles moderately developed; few, poorly-developed grayish supernumerary oval tubercles can be present on toes basis. Toes long, not webbed nor fringed, with ungual flaps present; relative length of toes, I<II<III<V<IV; toe IV average length 39.1% of foot length; toes I–V present disk expanded, rounded, dorsally emarginated, indented; disk pads even, broadened, following their disks sizes. The calcar tubercle may rarely appear poorly developed (e.g., CFBH 29055). Variation in size and proportions for adults are presented in Table 1. The adult male CFBH 29055 ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ) is herein used to illustrate new-collected specimens.
Color in life ( Fig. 5 View FIGURE 5 ). Despite of the general milky color of lectotype, which led Alípio de Miranda-Ribeiro to choose the epithet ‘lactea’ ( lacteus = milk), our results show that specimens of Ischnocnema lactea have stunningly attractive colorful patterns that display a large array of bright colors, mainly on hidden areas, with a high level variation on life coloration. The dorsum presents several blotches and/or large stripes that vary in position, dimension, shapes, colors, and intensity of tones, contrasting with background general color. An interocular blotch is usually present, being medially divided in some specimens; two small blotches between eyes and nostrils can be present, being connected as a stripe in some individuals; black supratympanic stripe conspicuous. The dorsum background color varies in brown, ranging from dark to light brown, nut-brown, orange, green, and grayish; some individuals also showed two or more colors on dorsal background. On flanks, colors fade into lighter variants of those colors, which may become cream nearby ventral surface. Forearms may exhibit two to three bars that can be faint or poorly conspicuous in some specimens; a blotch of varying color may be present on elbow. Thighs, shanks, and tarsus plus foot have three to four bars (each) that can be faint or inconspicuous in some specimens. Venter and hidden areas of inguinal region, thighs, shanks, and tarsus also present a large array of bright colors and patterns; background color ranged from intense to lighter tones of red, yellow, orange, black, gray, and cream/beige. Colors may be uniformly distributed all over the venter or varied in tones and intensity on different regions of undersurfaces, being, for example, lighter on gular and/or chest regions and more intense on belly and legs. The gular region on males can be darker than the rest of venter. The iris is golden with a medial red stripe crossing the black pupil, presenting several black vermiculations mainly in the ventral band. Two females, MZUSP 145318 and CFBH 25822, presented a colored dorsal band orange and green, respectively, externally lined by a dark line, delimiting dorsum from lateral surface.
Color in preservative. Background color in preservative faints into different tones of gray and silver. Our observations of old-dated fixed specimens revealed that hidden surfaces become slightly opaque, maintaining the same colors, while patterns and bright colors disappear with time. In general, all specimens maintained the former pattern of blotches, dots, and stripes in all surfaces.
Geographic distribution. Ischnocnema lactea is endemic to the Atlantic Rainforest biome, occurring in central portion of Serra do Mar, in the State of São Paulo, Southeastern Brazil. It inhabits mountainous forests at moderate altitudes, occurring from 700–1,120 m a.s.l. ( Fig. 6 View FIGURE 6 ). Coordinates of localities are provided in Appendix 2.
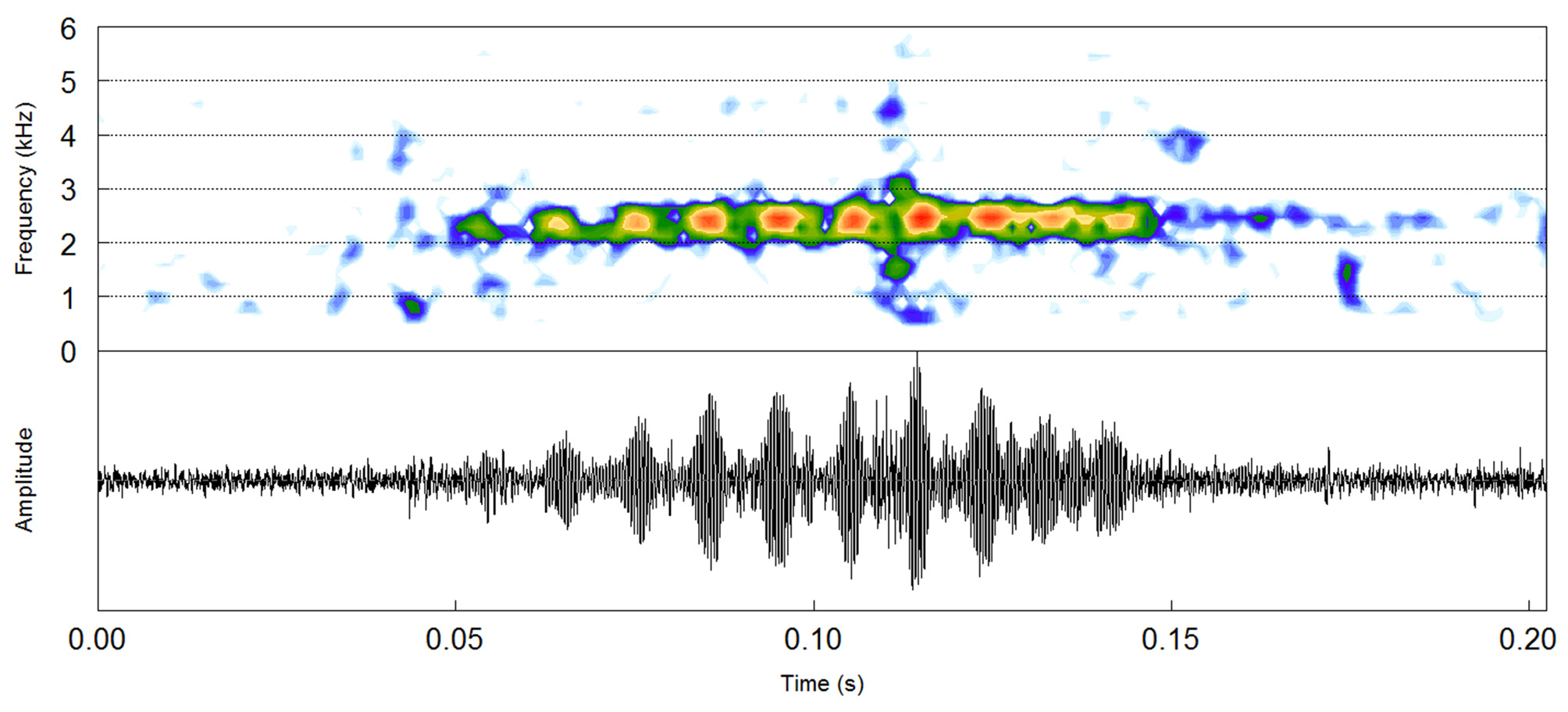
Vocalization and acoustic comparisons. The advertisement call of Ischnocnema lactea consists on a single pulsed note (notes emitted in series in I. nigriventris and I. randorum ), containing from eight to thirteen pulses ( Fig. 7 View FIGURE 7 ). The mean dominant frequency is 2418.9 ± 17.9 Hz (2239.5–2756.2 Hz, N = 24 notes). The mean note duration is 908 + 144 ms (634–1060, n = 24 notes). The mean of pulses per note is 10.45 + 1.10 (8–13, n = 24 notes), and note rate of 11.01 + 3.77 notes per minute (6.77–14.29, n = 24 notes). The mean of pulse duration is 6.5 + 1.2 ms (3.3–9.5 ms, n = 251 pulses). The mean of pulse rate is 113.83 + 12.45 s (94.33– 138.88 s, n = 24 pulses). The interval between pulses is evident mainly between the first and second pulses, on average 4.79 + 1 ms (3.53– 6.96 ms, n = 19 pulse intervals). The amplitude modulation between the other pulses is usually incomplete, i.e., there is no evident silence between them. The advertisement call of Ischnocnema lactea differs other species of its series by having pulses. Moreover, its call duration is longer than those of I. nigriventris and I. vizottoi , and shorter than that of I. randorum . Additional comparisons are provided in Table 2.
* For Ischnocnema lactea . call = note.
Natural History. The reproductive activity of I. lactea occurs during winter season (May-September) and appears to be associated with cold fronts that cause sudden drops in temperature along with rain, drizzle, and fog. Males call mainly during the day, starting early in the morning at around 07h00 a.m., decreasing the intensity at about 08h00 p.m. On one occasion, a male was recorded vocalizing until 09h00 p.m. Males vocalize perched on herbaceous vegetation, shrubs, and vertically on tree trunks in varying elevations (0.50–2 m, N = 8), and usually become darker when they are calling. Out of breeding season they become uncommon, and are rarely observed moving around on the leaf litter or resting on vegetation both during the day and at night. Two reproductive events were observed: the first occurred on 22 June 2010 at PNMNP (Vivian Trevine pers. comm.) during the arrival of a cold front, when ca. 200 males started calling in the morning from the litter (mainly) and shrub vegetation. Four to five pairs in amplexus ( Fig. 8 View FIGURE 8 ) were collected in pitfall traps. In the afternoon of the same day, the calling activity had already declined considerably, decreasing much more on 23th, the last day of the survey. The weather on 22 June, at 9:45 a.m. exhibited the following conditions: temperature 15.5o C, 84% humidity, cloudy, drizzling to light raining with fog; 23 June, at 8:30 a.m., temperature 14.4o C, 86% humidity, cloudy weather. The second event occurred on 15 June 2015 at the Núcleo Curucutu of PESM, in the municipality of São Paulo. About 150 males started vocalizing in the morning and continued through the afternoon, between 07h00 a.m. and 05:00 p.m., with a decrease of call activity after 04:00 p.m. The weather conditions consisted on a temperature varying between 12.1– 13.7°C, humidity 96–100%, cloudy with fog and drizzle.
Remarks. Our results based on morphologic data, demonstrating that several populations distributed throughout Serra do Mar belong to Ischnocnema lactea , are congruent with the results based on molecular data pointed by Canedo and Haddad (2012). According to the molecular phylogenetic hypothesis provided by the authors, which included data of some of the specimens analyzed herein (e.g., CFBH 14066 from Núcleo Santa Virginia, PESM, São Luis do Paraitinga; CFBH 16772 from Núcleo Curucutu, PESM, Itanhaém; and CFBH 29054 and 29107 from PNMNP, Santo André), these populations also belong to Ischnocnema lactea .
Furthermore, since the type locality of Ischnocnema lactea is no longer the municipality of Iguape, we propose a new English Name in exchange of the former ‘Iguape Robber Frog’ ( Frank & Ramus, 1995). Hence, considering its characteristic reproduction associated with sudden drops in temperature along with drizzle and fog derived from winter cold fronts, “Winter Drizzle Robber Frog” is the new English Name.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ischnocnema lactea ( Miranda-Ribeiro 1923 )
| Silva-Soares, Thiago, Malagoli, Leo Ramos, Cruz, Carlos Alberto Gonçalves & Caramaschi, Ulisses 2018 |
Basanitia lactea
| Miranda-Ribeiro 1923 : 851 |
Eleutherodactylus (Eleutherodactylus) lacteus
| Lynch & Duellman 1997 : 226 |
Ischnocnema lactea
| Hedges et al. 2008 : 27 |