
Echiothrix leucura Gray, 1867
|
publication ID |
https://doi.org/ 10.1206/871.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4627801 |
|
persistent identifier |
https://treatment.plazi.org/id/DE76879C-FF96-A304-0823-FAE15736A5AE |
|
treatment provided by |
Felipe |
|
scientific name |
Echiothrix leucura Gray, 1867 |
| status |
|
Echiothrix leucura Gray, 1867 View in CoL
HOLOTYPE: ‘‘No. 1499,’’ which is Gray’s catalog number. To learn more about this specimen, Musser queried Paula Jenkins at the Natural History Museum in London, who wrote (in litt., 1999) that the ‘‘specimen has apparently never been registered using the four-digit system, however, these Gray catalogue numbers are also traditionally accepted as register numbers.’’ ‘‘It is,’’ she continued, ‘‘listed in one of Gray’s catalogues … as: ‘1499 Skull a. the specimen fig. [the skull was illustrated in Gray, 1867: 599] Skin in a bottle in the Colln.’ There is no locality given here.’’ Paula also checked Thomas’
(1867: 599) original description.
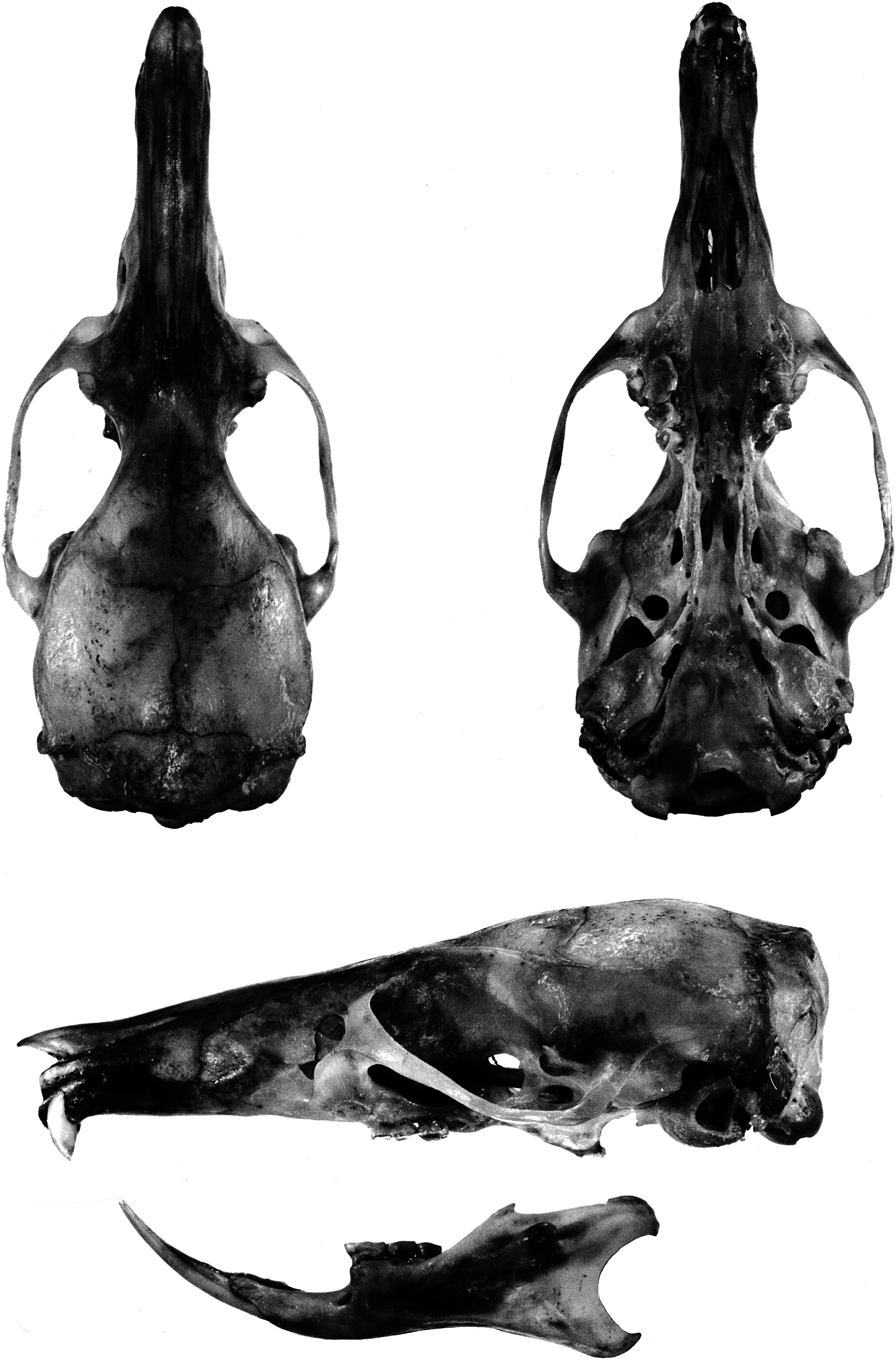
catalog and noted that ‘‘no number is given; the entry reads: ‘a. Sk. ad. male Australia. Type of genus and species.’’’ The identity of the collector was not recorded. Musser examined the holotype but at the time he took no measurements. The holotype is an adult male and consists of a dry skin and skull. Except for the damaged head, the skin is whole. The skull is incomplete ( fig. 3 View Fig ): anterior portion of the nasals, nearly all of the skull behind the orbit, and posterior half of each zygomatic arch are missing. The mandible is intact; upper and lower incisors and molars are present. The few measurements that could be made on the holotype are listed in table 3.
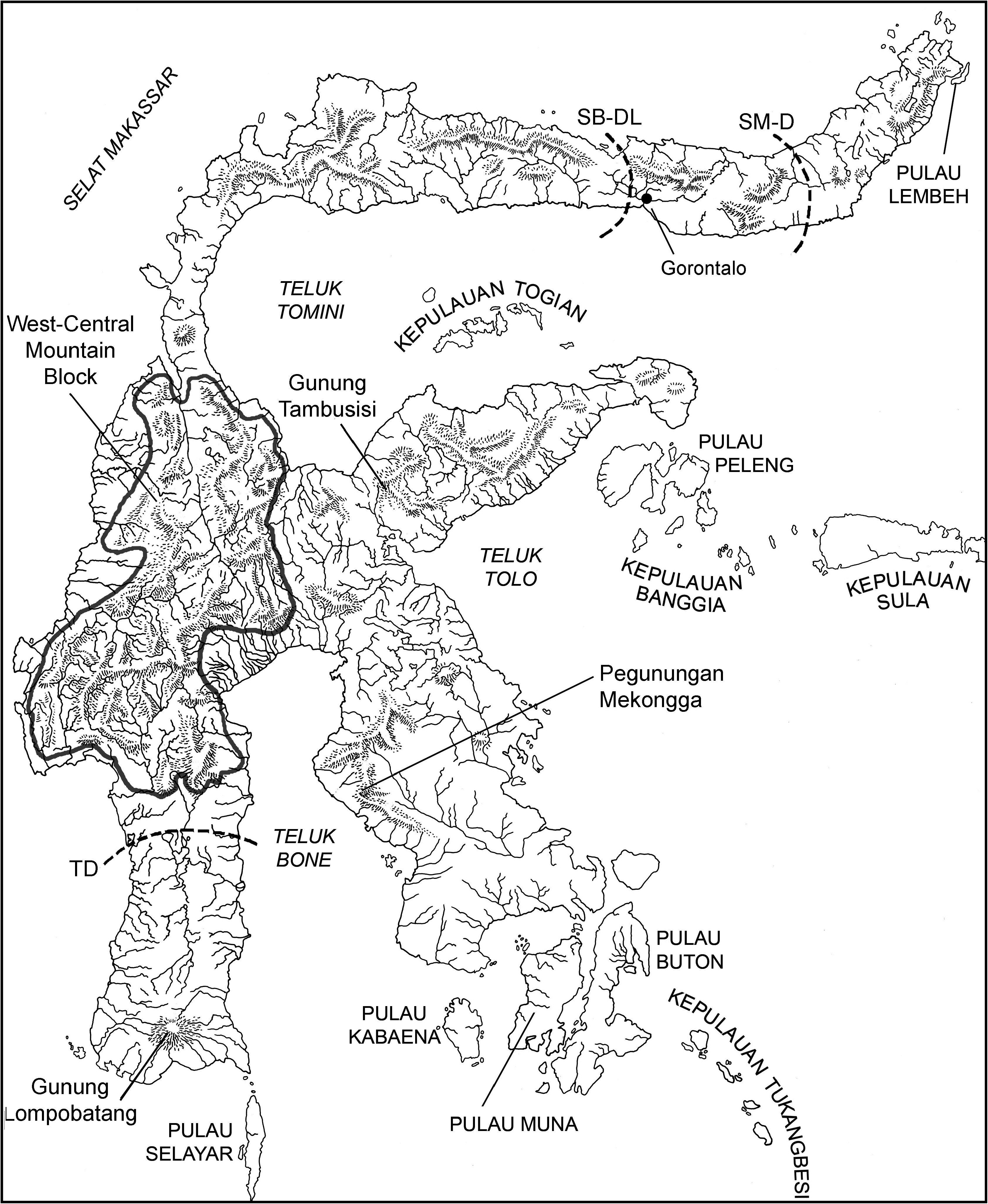
TYPE LOCALITY: Manado (01 ° 30 9 N, 124 ° 50 9 E; locality 1 in gazetteer and fig. 2 View Fig ), coastal plain near sea level, the northeastern tip of the northern peninsula of Sulawesi, Propinsi Sulawesi Utara, Indonesia GoogleMaps .
Gray (1867) indicated the specimen he described to have come from ‘‘ Australia.’’ No other reference to the actual provenance of the specimen exists. Paula Jenkins could not locate any additional information in the archives of the Natural History Museum (London). Subsequent researchers recognized that the holotype was not collected in Australia. Sixteen years after Gray’s description was published, Jentink (1883: 177) reported two additional specimens:
The type-specimen of this interesting species is in the British Museum and has been described by the late Gray as inhabiting Australia. A second specimen is in the Dresden Museum; it was brought from Menado, North-Celebes, some years ago. The specimen now before me is, as far as I know, the third representant of this species. I think it not improbable that Gray’s locality ( Australia) is incorrect and that later investigations will prove that it exclusively is an inhabitant of Celebes.
Tate (1936: 585–586) discussed specimens in the American Museum of Natural History that had been collected by G. Heinrich at Rurukan and noted that this locality is ‘‘at the extreme northeast of the Celebes and within a few miles of Menado, whence came the specimen in the Dresden Museum alluded to by Jentink,’’ and wrote that ‘‘In absence of evidence to the contrary the type locality of E. leucura may be restricted to Menado, north Celebes, making our series practically topotypical.’’
EMENDED DIAGNOSIS: Traits associated with external and cranial morphology as enumerated in the generic diagnosis also characterize Echiothrix leucura . The species differs from E. centrosa in possessing (1) a longer body and hind foot, but a shorter tail in relation to length of head and body; (2) a larger skull (as indexed by the greater mean values for occipitonasal and rostral lengths; interorbital breadth; height and breadth of braincase; and lengths of diastema, bony palate, and postpalatal region), but shorter incisive foramina and narrower rostrum,
TABLE 3 Age, Sex, and Measurements (mm) for Holotypes Associated with Species of Echiothrix a
bony palate, and mesopterygoid fossa; (3) larger molars and slightly more complex cusp patterns; (4) cusp t3 occurring at a higher frequency on second upper molar, a posterior cingulum typically present on first and second upper molars, (5) an anterolabial cusp present at a low frequency on second and third lower molars, posterior labial cusplets on all lower molars in some specimens, and a posterior cingulum present on first and second lower molars in most individuals surveyed.
SPECIMENS EXAMINED: Total 25 (see Gazetteer and Specimens).
ETYMOLOGY: The species name leucura is derived from the Greek leukos meaning ‘‘white.’’ Whether Gray meant to highlight the white ventral coat of Echiothrix or its white incisors (‘‘cutting-teeth white,’’ as Gray, 1867: 300, described them) is unknown, although we suspect the latter. Many species of murines have white underparts but not many possess incisors with white enamel layers.
GEOGRAPHIC AND ELEVATIONAL DISTRIBUTIONS: Voucher specimens indicate Echiothrix leucura is endemic to the northeastern end of the northern peninsula east of the Gorontalo region (00 ° 31 9 N, 123 ° 03 9 E; see the map in fig. 2 View Fig ). All specimens come from the mainland between coastal lowlands and approximately 1000 m (see gazetteer), an elevational interval that would be covered by tropical lowland evergreen rain forest GoogleMaps .
The geographic range of E. leucura is concordant with the distributions of several other mammalian species: the macaque Macaca nigra ( Fooden, 1969; Groves, 1980, 2005); and the murids, Bunomys fratrorum , Taeromys taerae , Rattus xanthurus , and R. marmosurus ( Musser and Carleton, 2005, unpublished MS.).
DESCRIPTION: Gray (1867: 600) provided a short description of E. leucura :
Fur dark grey brown, varied with black-tipped hairs on the back and sides; sides of nose, cheeks, throat, chest, and underside of limbs white; feet moderate, covered above with darkbrown hair; tail yellow, black at the base; cutting-teeth white; whiskers long, black, rather rigid. Length of body and head 9 K inches; tail imperfect; hind feet about 2 inches. Hab. Australia; British Museum, male? Tail imperfect.
His account does not do justice to the species. Echiothrix leucura is moderately large (table 4) with a lean body, long head, bristly dark gray or bluish gray upperparts, white underparts, mostly white tail that is longer than head and body, delicate front legs and small feet but robust hind legs with large and elongate hind feet, and very large ears ( fig. 4 View Fig ; Musser, 1990: fig. 3 View Fig ). The skull shows specializations such as a long and slender rostrum and a lack of pterygoid plates; the incisors are white or cream, and the molars are very small relative to skull size. The following description is drawn from adults; we have not seen examples of juvenile E. leucura , but their external characteristics are likely similar to juvenile E. centrosa , which are described in the account of that species.
Fur: The glossy dorsal coat of adults is harsh and bristly to the touch (not rigidly spiny as in hedgehog fur) and 15–20 mm long. It is composed of underhairs (or ‘‘wool’’ hairs; Voss, 1988) that form a thin underfur layer, and longer overhairs (also called ‘‘awns’’; Voss, 1988) comprising the overfur. The underhairs are soft, fine, wavy, and unpigmented along their entire lengths. The overfur layer consists mostly of wide and flat flexible spines, each with a channel extending along the dorsal surface, and a sharp tip. The base of each spine is unpigmented but the remainder is gray, which ranges in tone from pale gray near the unpigmented base to dark gray distally to the tip. In adults showing molt from old to new fur, the old spines have faded to straw brown, but those in the new layer proliferating through the skin are dark bluish gray. Scattered among the spines are softer thin hairs, each unpigmented except for a gray subterminal band. Guard hairs are inconspicuous because they are scattered throughout the coat and barely extend beyond the overfur layer; each is thin and round with an unpigmented base, long blackish subterminal band, and silvery tip. Overall, the fresh dorsal fur is dark gray or dark bluish gray frosted with white along the back but lightens to gray on sides of the head and body; old, worn pelage fades to pale straw brown.
The ventral coat of adults is 8–10 mm long on animals from low elevations but 10–13 mm thick over rats from higher places and consists of underhairs and overhairs but no guard hairs. Hairs forming the underfur are soft, fine, and wavy; those constituting the overfur are narrow, flat, and soft spines, each with a dorsal channel from base to near the tip. Texture of the coat is soft to slightly harsh to the touch but not spinous. Underhairs and overhairs are unpigmented in most specimens, so the ventral pelage, from tip of the muzzle to base of the tail, is overall white and sharply demarked from the gray dorsal coat. A few animals from Temboan show buffy or buffy orange patches on the chest and abdomen.
The white ventral coat is best appreciated in living rats or freshly prepared specimens. Some museum preparations, especially study skins stored since the early 1900s, appear pale yellow because the yellow dry skin shows through the fur or the hairs are stained with fatty oils (some of the skins from Temboan, for example, are obviously stained with yellowish deposits, so the entire animal appears to have a yellowish tint) and the underparts are yellow-orange (oily debris adheres to the hairs, and some skins are greasy yellow because subcutaneous fat was either not or incompletely removed before the skin was stuffed).
Fur covering the head of adults is pigmented like that clothing the body; throat and cheeks are white, chromatically indistinguishable from chest and abdomen. Rhinarium and lips are pink. Behind each ear is a crescent-shaped pale gray patch of soft, fine and short hairs that provides a whitish-gray crescent-shaped tuft around the base of each ear. The only obvious facial pattern is formed by blackish-brown eyelids and a circle of darker hairs around each eye, which on some individuals are more expansive and form a mask. All species have an array of mystacial, submental, superciliary, subocular, genal, and interramal vibrissae adorning the head; the longest mystacial extend beyond the ears when laid back over the head (see Brown, 1971, for descriptions of these sensory hairs and terminology). Some of the hairs lack pigment (appearing silvery), the rest are black or dark brown, and all have glossy surfaces.
Ears: Pinnae are very large relative to size of head and body (table 4). We have not seen living E. leucura , but the pinnae appear and feel rubbery in freshly caught E. centrosa , and while they seem naked, they are covered on inner and outer surfaces by short, fine, dark hairs that form a short and inconspicuous fringe along the dorsal internal rim of each pinna. Color of the ears is glossy dark gray with a pinkish tinge in living E. centrosa . We suspect texture and coloration to be similar in live E. leucura . Dried ears of stuffed museum skins lack the rubbery texture of the live animal and have dried to dark brown.
Tail: In samples of adults, average length of tail exceeds combined head and body length (LT/LHB 5 103.8 % –116.0 %; table 4). The tail is squarish in cross section and bicolored: in living animals, dorsal and lateral surfaces of the basal one-third to one-half range from dark gray to blackish gray, the rest of the tail, including the entire ventral surface, is glistening white (dorsal white length/tail length 5 47 % –67 %; table 5). In some material collected in the late 1800s and early 1900s, the pigmented basal portion of the tail has darkened to black and the white region altered to pale yellow, reflecting stain from the subcutaneous fat in the tail. Gray (1867: 600), for example, wrote of the holotype, ‘‘tail yellow, black at base.’’ White tail segments are yellowish in all of H.C. Raven’s material (stored at USNM), which was collected in the early 1900s. Slightly overlapping rings of moderately large scales (7–8 scale rings per cm, counted on the basal third of the tail) cover the tail. One, two, or three fine hairs (as
TABLE 5 Absolute Lengths (mm) of Dorsal White Tail Segment and Length of Tail, and Length of Dorsal White Segment
Relative to Length of Tail (%) In Samples of Echiothrix
Mean ± 1 SD and observed range in parentheses are listed.
long as one or two scales) emerge from beneath each scale. The tail appears to be scantily haired because the hairs are fine, inconspicuous, and vary in number per scale (instead of three at each scale, the usual pattern in murines), and the scutellation is exposed. Scale hairs are brown in the pigmented basal portion of the tail but unpigmented (silvery) in the white region.
Legs and feet: The front legs are short and slender, particularly the forearms. Each front foot has four slender digits ending in unpigmented stout claws and a tiny stubby thumb (pollex) bearing a nail ( fig. 4 View Fig ). The middle digits are the longest, the medial second digit shorter, and the lateral fifth digit extending about to the base of the adjacent digit. Dorsal surface of the metacarpal region and digits are white, covered with silvery hairs. Ungual tufts are sparse, consisting of one or two short hairs confined to the end of each digit, leaving the claws uncovered. Three interdigital pads along with a moderately large thenar and small hypothenar mounds form most of the naked palmar surface ( fig. 4 View Fig ), which ranges from unpigmented to dark gray (palmar surfaces of live E. centrosa have a rosy tinge reflecting the subcutaneous arterial circulation).
While the front claws are stout, moderately long, gently arched, and sharp, they are not excessively large relative to length of digits and size of the front foot ( fig. 4 View Fig ). By contrast, the montane Sulawesian shrew rats Melasmothrix naso , Tateomys rhinogradoides , and Paucidentomys vermidax , all consumers of earthworms, bear elongate curved claws that are very long relative to size of the foot ( Musser, 1982; Esselstyn et al., 2012).
The hind legs are large and robust. Hind feet are long and very narrow; the first (hallux) and fifth digits are much shorter than the three much longer middle digits, which are all about the same length ( fig. 4 View Fig ). The claw of the hallux barely reaches the base of the second digit, and the claw of the fifth digit extends to about middle of the second digit. Dorsal surfaces of the metatarsal region are either white (covered with unpigmented hairs), or white with small patches of gray speckling; digits are typically white. Conspicuous silvery ungual tufts cover bases of the ivory-colored stout claws. Each naked, dark gray plantar surface is adorned by six moderately fleshy pads: four interdigital mounds (forming a cluster at the bases of the digits), a tiny pimplelike hypothenar, and a short and narrow ridgelike thenar (both are very small and inconspicuous relative to plantar surface). The hind claws are curved and sharp tipped and moderately long relative to the length of the digits and hind foot, proportionally similar to the configuration seen in Sundaic Berylmys bowersii and Sundamys muelleri ( Musser and Newcomb, 1983: 15: fig. 15 View Fig ).
Teats: Two pairs of inguinal teats are characteristic of all female shrew rats surveyed (12 E. leucura , 15 E. centrosa ). Tate (1936) and Ellerman (1941) reported three pairs, which is incorrect.
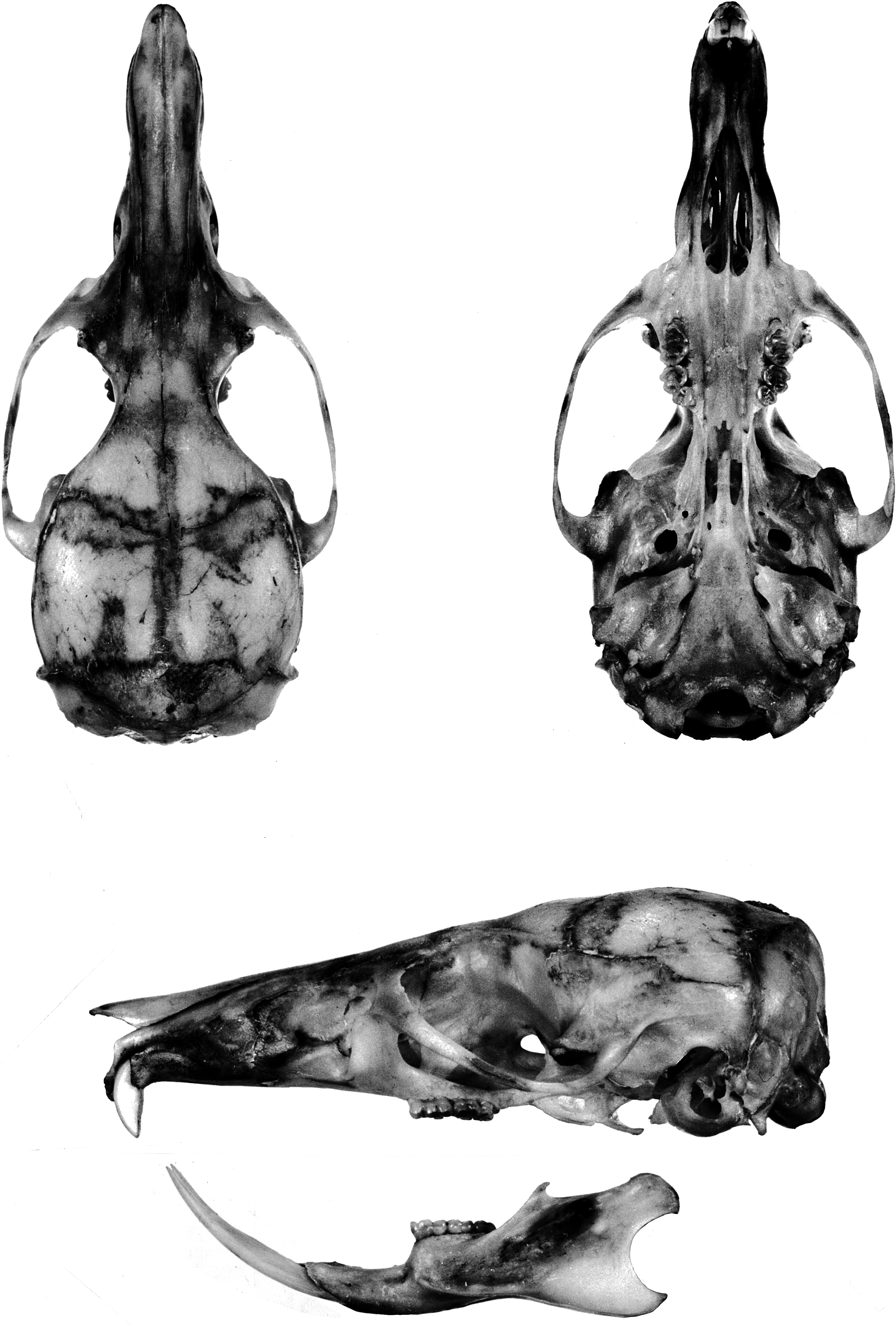
General cranial features: The cranial configuration of E. leucura is illustrated in the portrait of a skull from Rurukan ( fig. 5 View Fig ; the
6.6 mm), an adult female from Rurukan. X2.
5 6.0 mm), an adult female from Sungai Sadaunta. X2.
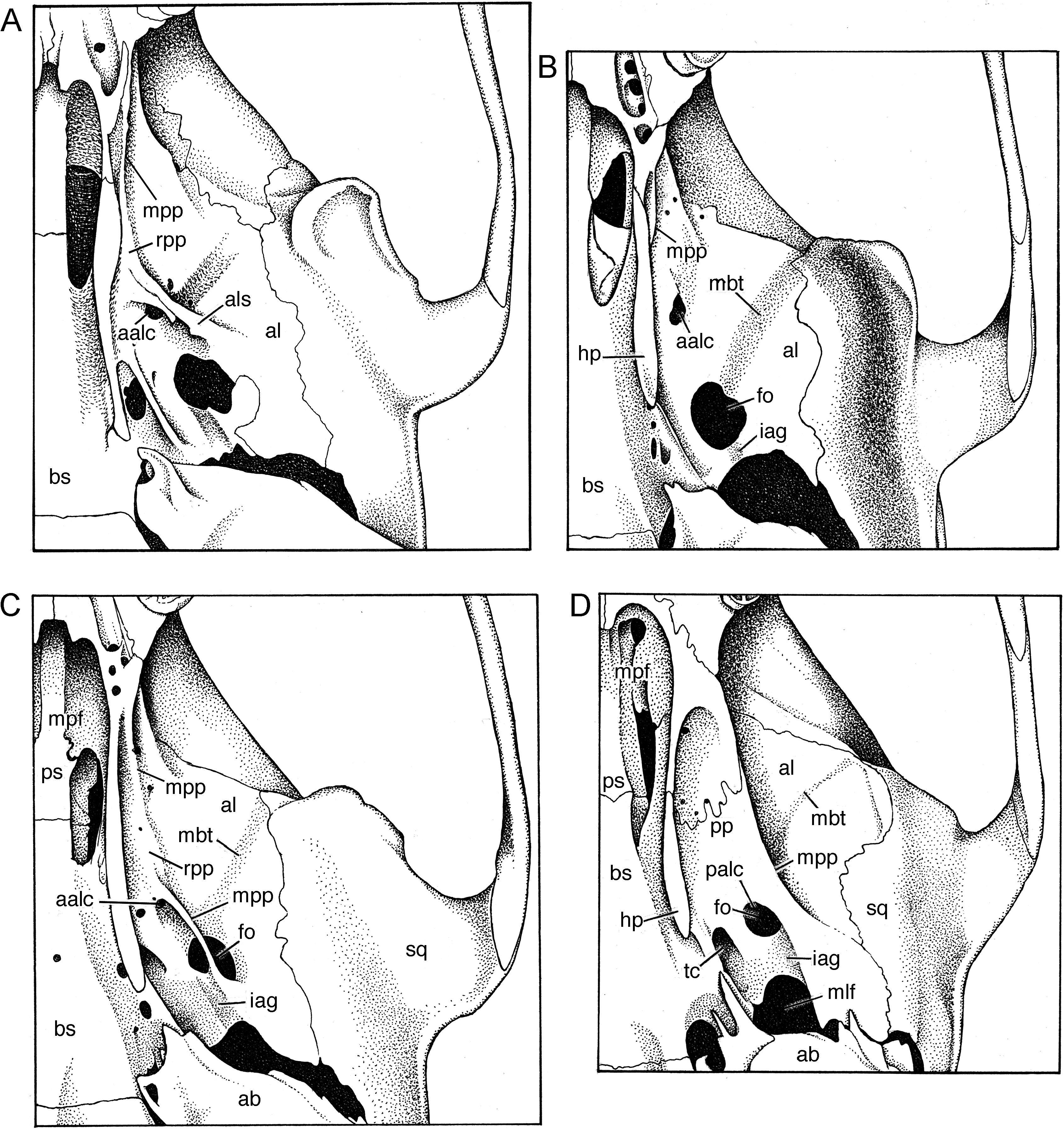
skull of E. centrosa from Sungai Sadaunta is portrayed in fig. 6 View Fig ). A long (about 40 % of occipitonasal length) and narrow (about 38 % of braincase breadth; table 6) rostrum is typical; from a lateral view, the rostrum tapers from its highest point level with the zygomatic plates distally to the incisors. The smooth sides are broken near the base of the rostrum by low nasolacrimal capsules that barely bulge beyond the rostral walls (and are hardly evident in dorsal or ventral views). Pointed tips of the nasals overhang the external nares and are either even with or project slightly beyond anterior edges of the premaxillaries; posterior borders of the nasals extend well beyond the premaxillary-frontal suture (by 14 % of nasal length). The zygomatic plate is moderately wide, its maxillary root originating posterior to the nasolacrimal bulge and above and in front of the first molar; its anterior margin is typically convex, is usually also inclined, and joins the dorsal maxillary root to form a conspicuous notch between the anterior edge of the plate and lateral side of the skull (as seen from dorsal perspective). The posterior edge of the zygomatic plate sits above the anterior onethird of the first molar. A tall and narrow infraorbital foramen is the usual configuration. The tendon of the superficial masseter muscle attaches to a small bony swelling or roughened raised area on the anteroventral corner of the zygomatic plate. Robust zygomatic arches bow appreciably outward; the maxillary and squamosal roots of each arch are united by a short jugal. The squamosal root of each zygomatic arch originates low on the outer braincase wall and its posterior margin extends as an indistinct ridge along the braincase to disappear well before the occiput.
A moderately wide interorbital region is usual. Its dorsolateral borders are defined by conspicuous ridges that extend along dorsolateral margins of the postorbital region (but never wide enough to form narrow shelves) and to the braincase where they form low but prominent temporal ridges that disappear anterior to the occiput (just before the lamboidal ridges). The otherwise smooth braincase is squarish (as seen from dorsal perspective) and deep (as viewed from the side) with a domed roof. A rectangular portion of the parietal drops below the dorsolateral margin of the braincase almost to the top of the zygomatic root; this projection and the squamosal form the wall of the braincase. The inner sidewalls of the braincase are smooth, without squamosalalisphenoid grooves. Sides of the braincase are vertical from the temporal beading to squamosal roots of the zygomatic arches. The occipital region is moderately deep and roofed by the interparietal in the middle and dorsal segment of the exoccipital on either side. The boundary between squamosal and exoccipital is marked by prominent lamboidal ridges. The posterior wall of the occiput is vertical (in lateral view) and even with the posterior boundaries of the occipital condyles. The squamosal above each auditory (ectotympanic or bullar) capsule and just anterior to the lamboidal ridge is complete (not penetrated by a subsquamosal foramen).
As seen from the ventral perspective, the moderately long incisive foramina (about 50 % of diastemal length; table 6) are located in the center of the diastema and form an elongate teardrop in outline. Just behind the upper incisors is a slitlike interpremaxillary foramen. Except for a pair of grooves that increase in depth from front to back, the bony palate is smooth, its posterior margin projects well past the third molars to form a bony shelf. A pair of large posterior palatine foramina penetrate the palate at the maxillopalatine suture opposite either first or second upper molars. Maxillary molar rows are either parallel or bow medially toward the back of the bony palate. The mesopterygoid fossa is narrow and its dorsolateral walls are perforated by two wide and moderately long sphenopalatine vacuities that expose the medial borders of the presphenoid and basisphenoid.
Each ectotympanic bulla (5 bullar capsule) is moderate in size (about 11 % of the occipitonasal length) and bears a wide and long bony eustachian tube. The medial sagittal plane of each bullar capsule is oriented ventromedially, so the capsules appear to rest on the basicranium and project toward the midline rather than more nearly vertical (an orientation similar to that seen in the Philippine Tarsomys apoensis [ Musser and Heaney, 1992: 23, fig. 9 View Fig ]). The bullar capsule does not cover the entire surface of the enclosed periotic bone, leaving exposed a thick posteromedial wedge and a narrow flange extending forward between the ectotympanic and basioccipital. The carotid canal is bounded by part of the periotic, adjacent ectotympanic, and lateral border of the basioccipital (pattern is closely similar to that illustrated for Philippine Chrotomys [ Musser and Heaney, 1992: 78, fig. 43]). All specimens possess a large stapedial foramen penetrating the crevice (the petromastoid fissure) between the bullar capsule and the periotic. A middle lacerate foramen, either spacious or narrow, separates the bullar capsule from the posterior margin of the alisphenoid.
In lateral view, a narrow flange of periotic is exposed along the anterodorsal margin of the bullar capsule. The capsule and periotic are separated from most of the squamosal by a broad postglenoid foramen that is confluent with a ventral middle lacerate foramen. The mastoid portion of the periotic is slightly inflated, its outer surface without perforations.
Within the orbit, the ethmoid foramen is small and the optic foramen large, a typical murine pattern. Orbitosphenoid, alisphenoid, and frontal bones join to form a solid section of the braincase wall, unbroken by a sphenofrontal foramen. Sphenopalatine and dorsal palatine foramina are separate, a pattern similar to that found in species of Rattus ( Musser, 1982: 22) .
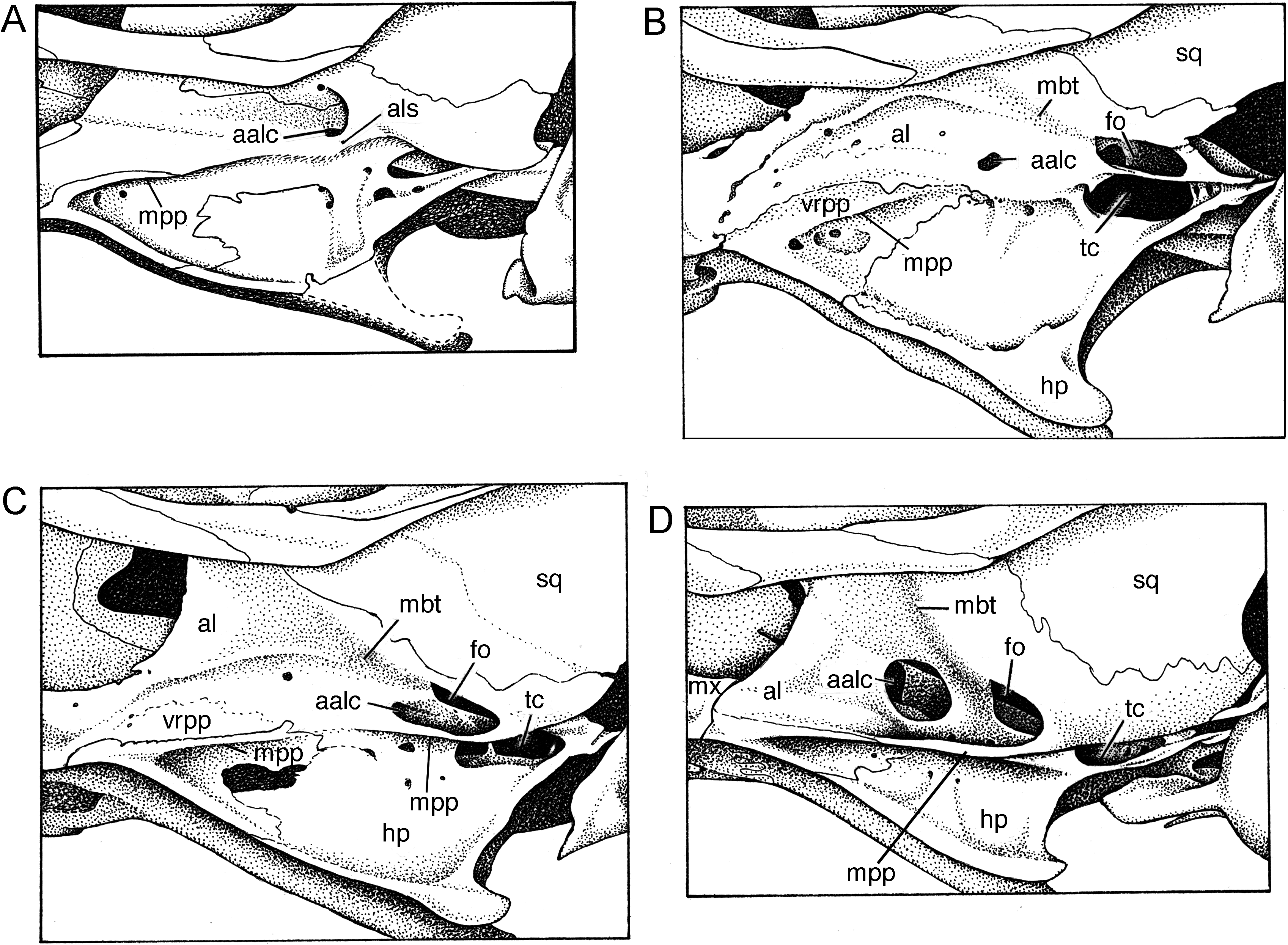
Pterygoid region: Osseous structure of the pterygoid region of Echiothrix is extremely simplified: the pterygoid plates have regressed and are represented only by diminutive remnants. Among murines, this osteological design of the pterygoid-alisphenoid region is repeated only in the Philippine shrew rat Rhynchomys (see the cranial illustrations in Musser and Heaney [1992: 78] and Balete et al. [2007: 293]) and the Sulawesian shrew rat Paucidentomys (Esselstyn, et al., 2012). The best way to appreciate this severe modification is to first describe the pterygoid region in the Sulawesian Maxomys dollmani , which expresses a pattern common to nearly all other murines, especially those from the Indomalayan region, Sulawesi, Philippines, New Guinea, and Australia (see the cranial illustrations in Musser, 1982, 1991; Musser and Newcomb, 1983; Musser and Holden, 1991; Musser and Heaney, 1992; Flannery, 1995; Musser et al., 2008; Musser and Lunde, 2009; Heaney et al., 2012; Balete et al., 2012), as well as species in Europe and Africa (see the cranial drawings in Happold, 2013).
In Maxomys dollmani , each pterygoid plate forms a long and wide shelf bounded on the medial side by a hamular process and defined laterally by a smooth margin; the palatine bone forms the plate’s anterior half and the pterygoid bone its posterior section ( fig. 7D View Fig ). Nearly all of the ventral surface of the pterygoid plate anterior to the foramen ovale is shallowly excavated and forms the pterygoid fossa. The posterolateral and posterior edges of the plate converge behind the foramen ovale to form a wide and smoothly rounded ridge, which defines the anterolateral border of the spacious medial lacerate foramen separating the pterygoid plate and posterior margin of the alisphenoid from the ectotympanic bullar capsule. Just medial to this pterygoid ridge is a deep groove for the infraorbital branch of the stapedial artery. The spot where the artery leaves the groove and passes to the dorsal surface of the pterygoid plate defines the posterior opening of the alisphenoid canal. Medial to the foramen ovale is the sizeable opening of the transverse canal.
As seen from lateral perspective ( fig. 8D View Fig ), a bony alisphenoid strut is not present in Maxomys dollmani , a loss resulting in the coalescence of the foramen ovale accessorius and masticatory-buccinator foramina (see fig. 48 in Musser and Newcomb, 1983: 457, showing the configuration when an alisphenoid strut is present). Exposed to view is the anterior opening of the alisphenoid canal, the open canal itself, the foramen ovale and the defining lateral margin of the pterygoid plate. Emerging from the foramen ovale is a shallow trough for the masticatory and buccinator divisions of the maxillary nerve.
In Echiothrix , the pterygoid plates have disappeared as major horizontal structural entities and only remnants remain, as seen from ventral perspective of skulls ( fig. 7 View Fig A– C). In all three views of E. centrosa , most of the alisphenoid bone extends to the base of the hamular process without meeting a generous horizontal pterygoid plate (panel A) and the entire outer surface of the alisphenoid is exposed. Aside from tiny scattered nutrient foramina, only two major openings are present in the alisphenoid: the anterior opening of the alisphenoid canal forms a small round or irregular aperture anterior to the much larger and spacious foramen ovale. Emerging from the latter is a shallow trough for the masticatory and buccinator divisions of the maxillary nerve. Between the foramen ovale and the large middle lacerate foramen is a shallow groove in which the infraorbital artery courses. In some specimens, a continuation of the groove can be detected between the anterior rim of the foramen ovale and the anterior opening of the alisphenoid canal ( fig. 7C View Fig ) but in others the groove exists only between the middle lacerate foramen and the foramen ovale ( fig. 7 View Fig A–B). Apparently the infraorbital artery courses along the surface of the alisphenoid in Echiothrix but in Maxomys is concealed by the pterygoid plate and is exposed only between the middle lacerate foramen and foramen ovale, the latter marking the posterior opening of the alisphenoid canal.
Variation in vestiges of the pterygoid plate is also shown in figure 7 View Fig . The skull in panel A retains an alisphenoid strut to which a sliver of pterygoid plate margin is attached; forward of the strut is a slight ridgelike remnant of the anterior margin of the plate. Panel B reflects the configuration of the alisphenoid region in many examples of Echiothrix where a short sliver of the anterior plate margin is the only sign of the pterygoid plate from ventral perspective. The skull depicted in panel C preserves a narrow ridgelike relic of the anterior plate margin and a slim crescent of bone representing the posterolateral margin of the pterygoid plate; stippling that connects the anterior part of the bony crescent with the relict anterior plate margin outlines a narrow ledge (remnant of the pterygoid plate) undefined by a discrete lateral margin; a comparable narrow ledge or platform is found in many examples of Echiothrix .
The alisphenoid region, as seen from lateral perspective, of the same three specimens of E. centrosa that are illustrated in figure 7 View Fig is shown on panels A–C in figure 8 View Fig . In panel A, a bit of the pterygoid plate margin is evident as is an alisphenoid strut, which is typically absent in most examples of Echiothrix ). The anterior opening of the alisphenoid canal is small (also seen in panels B and C), in contrast to the comparable spacious opening in Maxomys (panel D). Vestiges of the pterygoid plate margin are also evident in panels B and C. So is the encroachment of what appears to be a low vertical remnant of the pterygoid plate into the alisphenoid. This vertical remnant apparently represents the transformation of a horizontal pterygoid ridge—the usual murine conformation—into a vertical surface that, along with the lower portion of the alisphenoid, forms the outer wall of the mesopterygoid fossa (mpf in fig. 7 View Fig ); this vertical remnant varies in extent among specimens of Echiothrix , ranging from the expanse illustrated in panels B and C to a thin sliver just above the ridgelike pterygoid margin. In most species of murines, such as Maxomys dollmani (panel D), the alisphenoid extends from its suture with the squamosal ventrally to the base of the pterygoid plate and anteriorly to the maxillary (mx) just behind the molar row. The outer wall of the mesopterygoid fossa is formed only by the alisphenoid.
Cephalic arterial pattern: All specimens of Echiothrix leucura (and E. centrosa ) surveyed possess a carotid arterial plan that is primitive for members of the subfamily Murinae (character state 2 of Carleton, 1980; pattern 2 described by Voss, 1988; conformation diagrammed for Oligoryzomys by Carleton and Musser, 1989), but derived for muroid rodents in general. The pattern is reflected in certain cranial foramina and osseous landmarks seen in cleaned skulls. No sphenofrontal foramen penetrates the bony junction of orbitosphenoid, alisphenoid, and frontal bones; no squamosal-alisphenoid groove scores the inner surface of each wall of the braincase; and the stapedial foramina is neither minute nor absent. Instead, there is a large stapedial foramen in the petromastoid fissure, and a shallow groove (iag in fig. 7 View Fig B– C) extending from the middle lacerate foramen (mlf in fig. 7D View Fig ) to the foramen ovale (fo in fig. 7 View Fig B–D). This disposition of foramina and grooves indicates that the stapedial artery branches from the common carotid and enters the periotic region through a large stapedial foramen. The infraorbital branch of the stapedial artery exits the pteriotic through the middle lacerate foramen, courses in a short groove (between middle lacerate foramen and foramen ovale), and continues on the outside of the skull at the base of the hamular process to disappear into the braincase through the anterior opening of the alisphenoid canal from which it emerges to course through the anterior alar fissure into the orbit. The supraorbital branch of the stapedial is absent; the arterial supply to the orbit is furnished by the distal part of the infraorbital branch. This circulatory plan is common among murines ( Musser and Newcomb, 1983; Musser and Heaney, 1992) and is also found in some North and South American cricetids ( Carleton, 1980; Voss, 1988). The derived murine version of the carotid arterial supply is contrasted with the primitive configuration and another more derived pattern, both of which are found within other muroid rodents as documented by Bugge (1970), Carleton (1980), Carleton and Musser (1989), and Voss (1988).
Mandible: Each gracile dentary is elongate, particularly the diastemal portion between the incisor and anterior margin of the first molar ( fig. 5 View Fig ). On the dorsal rim of the ramus behind the molar row, the coronoid process is a minute triangular projection, and the long rim between it and the large and long condyloid (articular) process is either straight or slightly concave (very shallow sigmoid notch). The posterior margin of the dentary between the large condyloid and angular processes is deeply concave (outlining a half circle). A large bulge on the side of the dentary behind the coronoid process and even with the dorsal rim marks the posterior termination of the incisor sheath (alveolus). Masseteric ridges on the lateral surface of each dentary are pronounced.
Dentition: Upper incisors are small, short, and narrow, and emerge from the rostrum at a right angle (orthodont in form; see Thomas [1919] and Hershkovitz [1962: 103] for definitions of incisor configurations). The anterior face of each incisor is scored by two shallow grooves ( fig. 3 View Fig ). Dentine is white (unpigmented) as is the enamel in most specimens, but a few show a pale yellow tinge in the enamel layers. In live E. centrosa , the teeth appear ivory and emit a translucent quality; we suspect the teeth appear similar in live E. leucura . Enamel layers form the face and anterolateral third of each incisor, and dentine the remainder, a configuration similar to that found in most murines (as illustrated for Rattus in Musser and Heaney, 1992: 79).
Beyond their emergence from the anterior body of the dentary, the lower incisors are awl shaped—each is long, slim, gently curved, and with a sharp tip and elongate wear (occlusal) facet; enamel layers are smooth, without grooves. While the dentine is white, the enamel layers range from white to a pale yellow tinge—usually the distal portion of each incisor is unpigmented but the basal portion pale yellow. The extent of the enamel layer is similar to the pattern seen in most murines, Rattus , for example ( Musser and Heaney, 1992: 79, fig. 46). The full length of each long incisor is slender and although gracile in form the tooth is not weak. Twothirds of the incisor is contained in a sheath within the body of the dentary where it ends between coronoid and condyloid processes in an elongate bulge on the lateral surface of the dentary. This bony enclosure provides the incisor with foundational strength for stabbing prey (see Natural History Particulars of E. centrosa ).
Every maxillary molar is secured by three roots. The alveolar patterns illustrated for Maxomys wattsi (Musser, 1991: 32, fig. 18 View Fig ) are also usual for Echiothrix . Each first and second molar is anchored by large anterior and lingual roots along with a smaller posterior holdfast; each third molar is held in place by three roots of about the same size. There are two large roots beneath each of the mandibular molars.
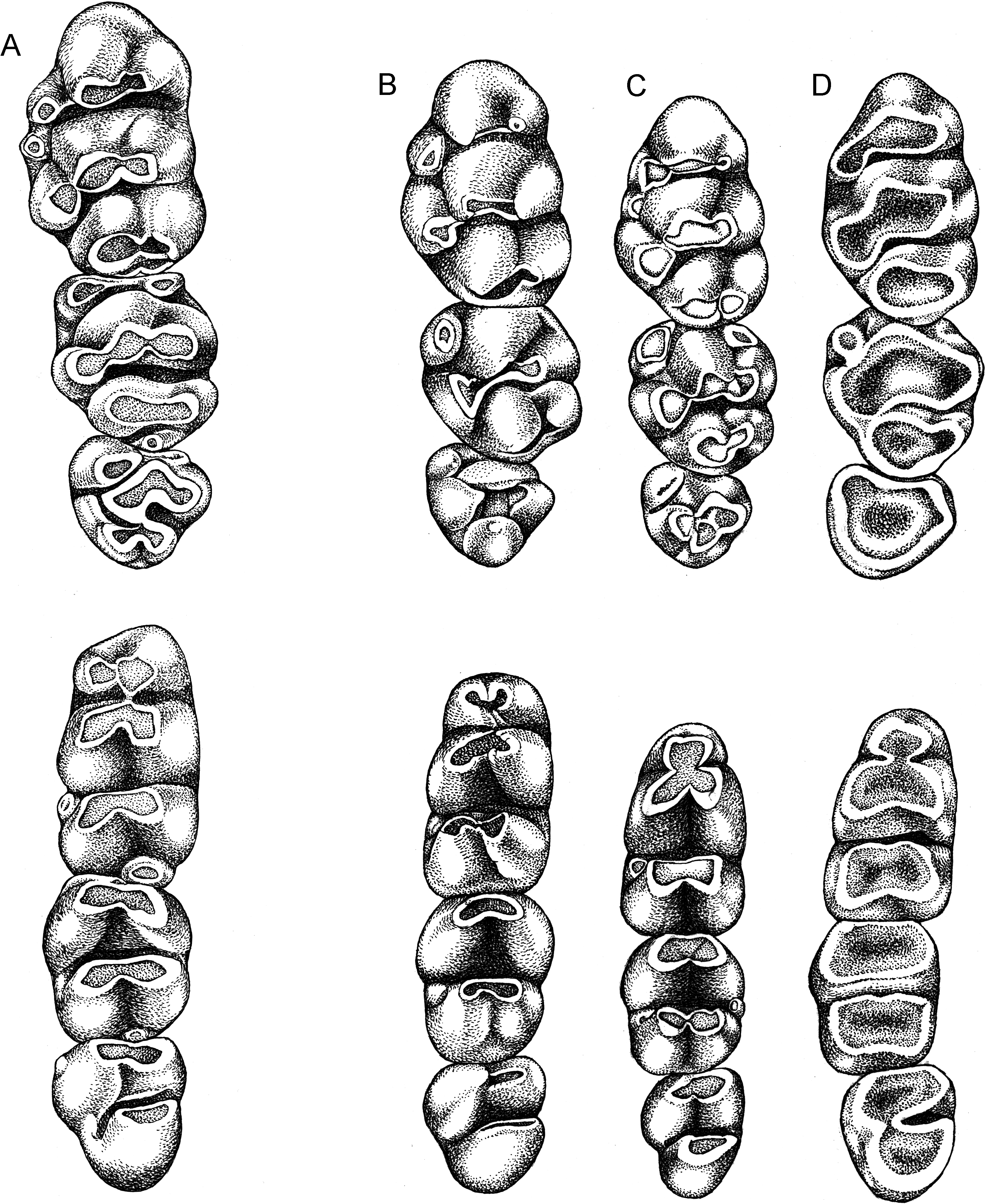
The brachydont molars are small and molar rows are short relative to size of skull (13 % of occipitonasal length; table 8); the rows are either parallel or curve gently posteriorly toward the midline of the bony palate. Molars grade in size within each row: the first is the largest, the third the smallest, which is the usual configuration observed in most species of murine rodents ( Carleton and Musser, 1984). Like many murines, the cusp rows incline caudad, so that within each maxillary row the first molar overlaps the second and that tooth leans slightly against the third; the third molar in each mandibular row inclines against the second, which slightly overlaps the first molar. The rows of cusps on each tooth are moderately close to one another, a condition usually associated with inclined rows of cusps; in murines with widely separated rows, the cusps are typically erect.
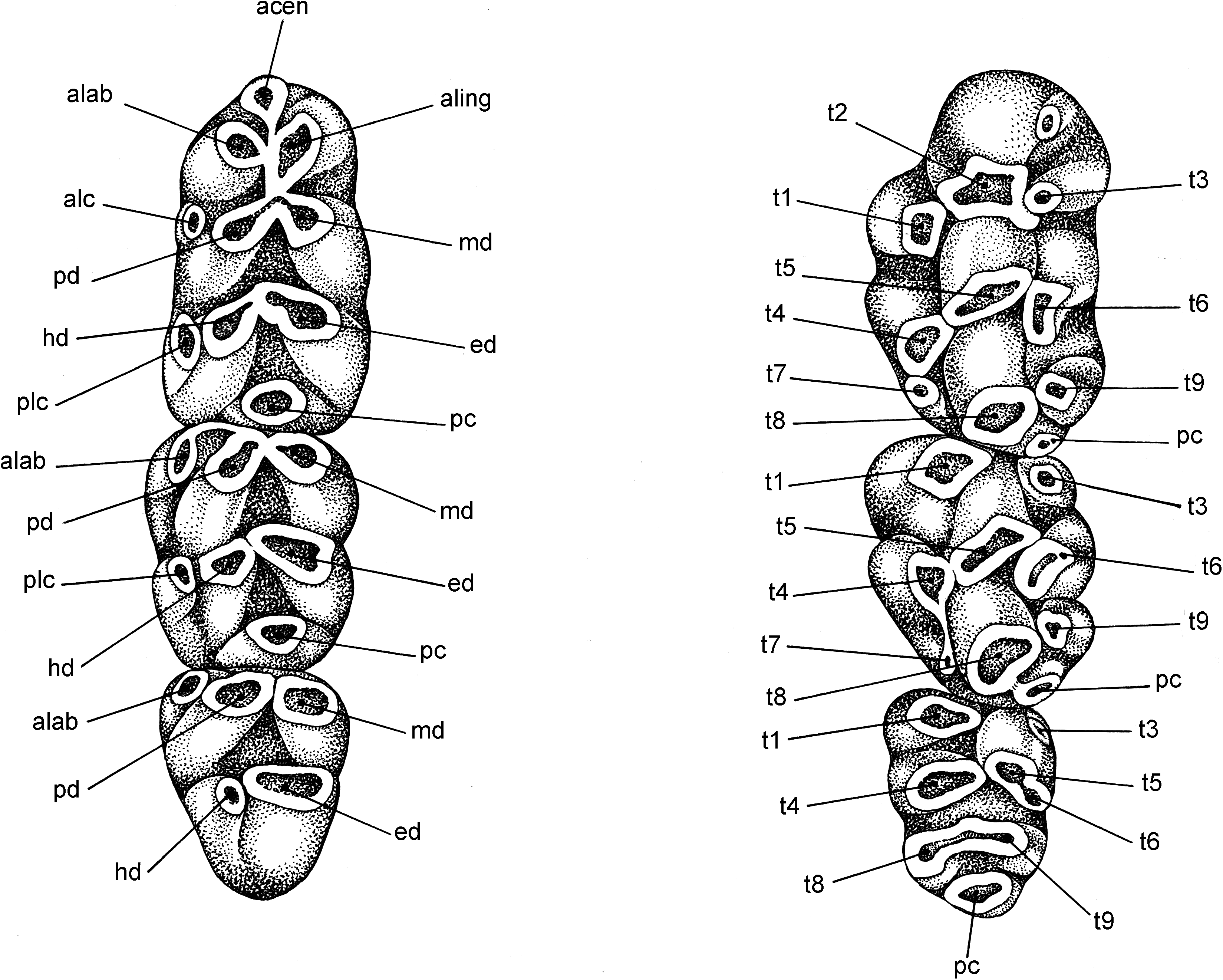
Occlusal surfaces of maxillary molars are formed by simple rows of low cusps and lack the complexities found in more complicated patterns that characterize some other murines ( fig. 10 View Fig , upper row). For example, there is no cusp t7, which is a prominent feature on the occlusal surface in certain murine genera ( Lenothrix , for example; fig. 9 View Fig ); cusp rows stand free, unconnected by labial or lingual enamel bridges (stephanodont crests as described by Misonne, 1969: 55); although close to one another, cusps t4 and t8 do not coalesce along their lingual margins until they are worn to the cingulum; the anterior cingular face on each first molar is smooth, without a shelflike ridge or small cusp (a cingular ridge, often bearing a small cusp, is a usual element on the first molar in Rattus hoffmanni , for example; Musser and Holden, 1991: 348). An occasional specimen of E. leucura shows an auxiliary cusp behind cusp t1 on the first molar ( fig. 10A View Fig ).
On the first upper molar, the labial and central cusps in each row are transversely aligned; the lingual cusps t1 and t4 are situated well back of the first and second rows, respectively. A posterior cingulum occurs on nine of the 10 specimens surveyed (sample size is small because teeth are missing from some specimens or in others the occlusal surfaces are too worn to identify a posterior cingulum). The posterior cingulum extends from the back of cusp t 8 in the form of a ridge, a triangular projection, or a large cusp projecting labially to form a portion of the occlusal surface (as exemplified by Lenothrix ; fig. 9 View Fig ).
On the second molar, the first row of cusps is represented by cusp t1, which is always present, and cusp t3, which is present in only about half the sample surveyed. A posterior cingulum was located on nine of 10 specimens.
Cusp t1 is present on the small third molar, but cusp t3 is not. Cusps 4, 5, and 6 form a structure that is usually chevron shaped. Cusps 8 and 9 form a posterior lamina in some specimens, but in others only a single structure is present, representing either cusp t8 only or completely merged cusps t8 and t9; the third molar lacks a posterior cingulum.
Occlusal topography of each mandibular molar consists primarily of chunky transverse rows, each representing the fusion of two cusps ( fig. 10 View Fig , lower row). A large posterior cingulum, generally elliptical in cross section, occurs on the first lower molar in 11 of the 12 specimens surveyed, and a much smaller posterior cingulum could be identified on the second molar in seven of 10 specimens. Located at the front of the first molar is a chunky anteroconid composed of large anterolabial and anterolingual cusps (no anterocentral cusp was found on any of the specimens) that have merged to form a large oblong lamina (without discernable cusp boundaries in some specimens, but clearly formed from two cusps in others) either slightly or much narrower than the lamina behind it. None of the molars bear anterior labial cusplets; various frequencies of occurrence of posterior labial cusplets on all molars along with anterolabial cusps on the first and second molars comprise minor components of the occlusal surface.
COMPARISONS: Cranial and dental morphometrics as well as qualitative cusp differences and geographic distributions between E. leucura and E. centrosa will be offered in the account of the latter species.
GEOGRAPHIC VARIATION: The samples of E. leucura used in analyses of morphometric traits come from four places and each analytical sample is small. There is insufficient material and geographic coverage for any serious analysis of geographic variation in external, cranial, and dental traits. We can report that features of fur coloration along with lengths of head and body, tail, hind foot, and ear among adults show hardly any differences from place to place other than that ascribed to individual, age, and sexual variation. Nor do definable groups emerge when cranial and dental measurements are subjected to principal-components analysis. The distribution of specimen scores for the samples from Rurukan, Gunung Masarang, Tondano, and Temboan projected on the first and second principal components (not illustrated here) intermix to form a single large cluster. If significant geographic variation exists within the range of E. leucura , it will have to be demonstrated in the future by obtaining larger samples from more places and adding analyses of DNA sequences to results derived from morphometric inquiry.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.