
Diomini Gordon, 1999
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4554.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:A804E949-109A-468D-B58B-CF7C8BCB3059 |
|
DOI |
https://doi.org/10.5281/zenodo.5921864 |
|
persistent identifier |
https://treatment.plazi.org/id/DF18F971-FFC7-0B2C-FF4B-FA206D1BBE47 |
|
treatment provided by |
Plazi |
|
scientific name |
Diomini Gordon, 1999 |
| status |
|
Tribe Diomini Gordon, 1999
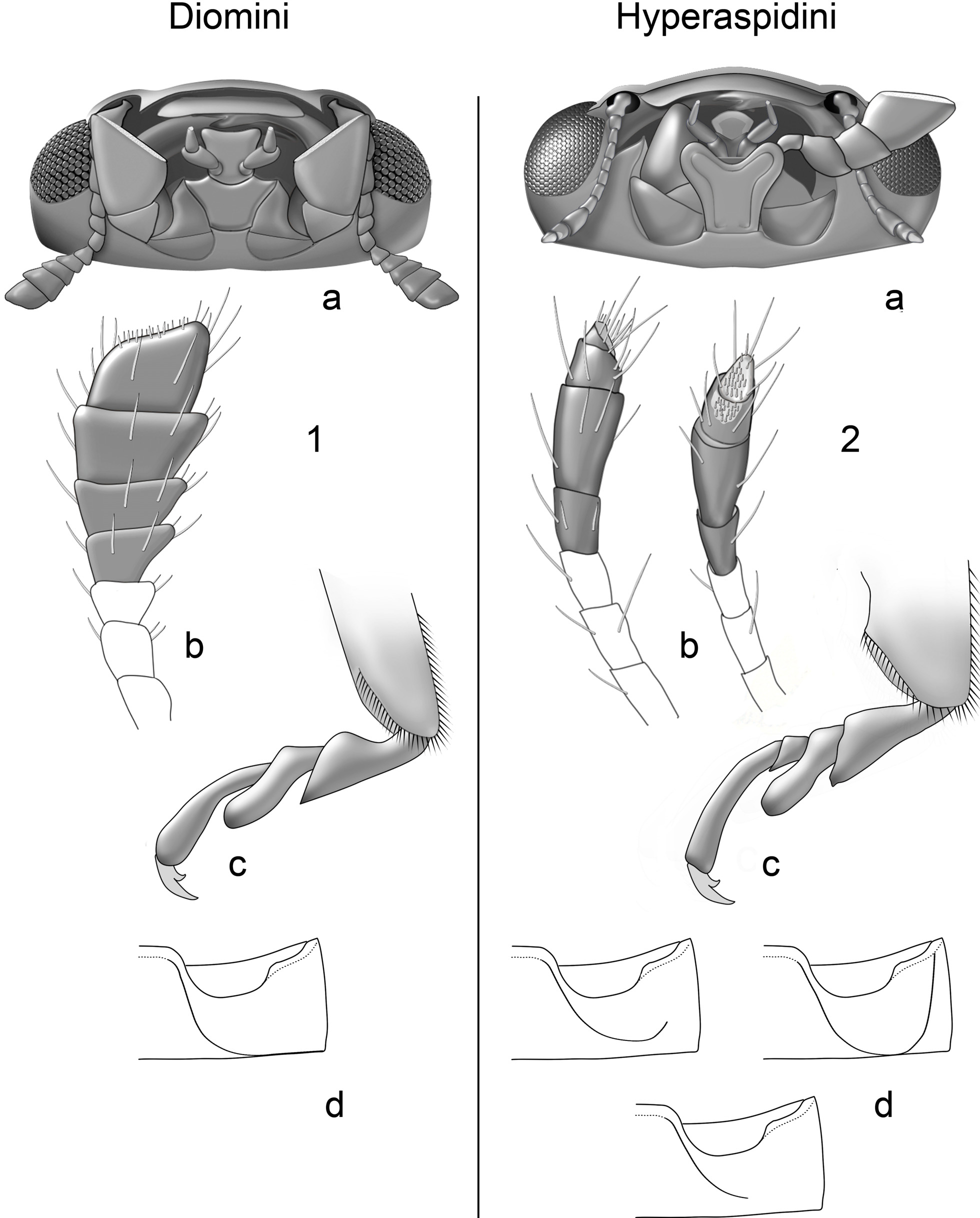
( Figs. 1 View FIGURES 1–2 a–d)
Diomini Gordon 1999:3 . Type genus: Diomus Mulsant, 1850 Included taxa (new circumscription, excluding Magnodiomus and Erratodiomus which we transfer to Hyperaspidini ; see “ Remarks,” below): Diomus, Decadiomus View in CoL , Heterodiomus , Dichaina , Andrzej , and the new genus described herein
Diagnosis. Size minute to small (1.1–3.5 mm), pubescent; antenna ( Figs. 1a,b View FIGURES 1–2 ) composed of 10 or 11 antennomeres; pedicel bead like, articulated with and slightly narrower than scape, antennomere 3 elongate, 1.5–3× as long as antennomere 4; distal 3, 4, or 5 antennomeres forming slightly flattened asymmetrical compact club with oblique to truncate apex bearing concentration of short sensory setae (=sensilla). Mentum ( Fig. 1a View FIGURES 1–2 ) subtrapezoidal, tapered posteriorly, typically with anterior margin concave or bicuspidate. Terminal maxillary palpomere in repose free, not partially inserted beneath mentum, more or less expanded distally with sensory surface directed anteriorly or anteromedially. Tarsi trimerous ( Fig. 1c View FIGURES 1–2 ). Tibial spurs lacking. Abdomen with 6 visible ventrites; ventrite 1 and 2 partially fused medially. Abdominal postcoxal line ( Fig. 1d View FIGURES 1–2 ) curving posterolaterally, merging with posterior margin of ventrite. Male genitalia with basal lobe (=penis guide sens. Ślipiński) distinctly asymmetrical to roughly symmetrical at least in outline; penis capsule with outer arm distinct to obsolete ( Figs. 9–12 View FIGURES 3–12 ). Female genitalia with spermathecal capsule well developed, except vestigial or absent in some Australian species; capsule with simple angular or C-shaped form, vermiform throughout or with swollen basal chamber; cornu of moderate length, not convoluted; ramus and nodulus sessile to weakly projecting; ramus with or without short beak-like projection (apodeme) overhanging attachment of accessory gland. Coxites teardrop or rhombus-shaped, with widely arcuate posterior margin.
Members of Diomini are most easily confused with other small pubescent lady beetles. The shape of the abdominal postcoxal line and wide posterior margin of the coxites will distinguish diomines from both Scymnus and Nephus (including its subgenera sens. Gordon 1985 which are now often accorded full generic status). In Scymnus and Nephus the postcoxal line does not reach the hind margin of the ventrite and the outer end is often recurved toward the base of the ventrite; the coxites are narrow and distally tapered rather than broad. Among New World species, the basal lobe of Diomini is distinctly asymmetrical, while that of Scymnus is bilaterally symmetrical. Furthermore, Scymnus can be distinguished from Diomini by the possession of cryptotetramerous tarsi, and Nephus is distinct in having the basal two antennomeres fused or tightly joined (non-articulated).
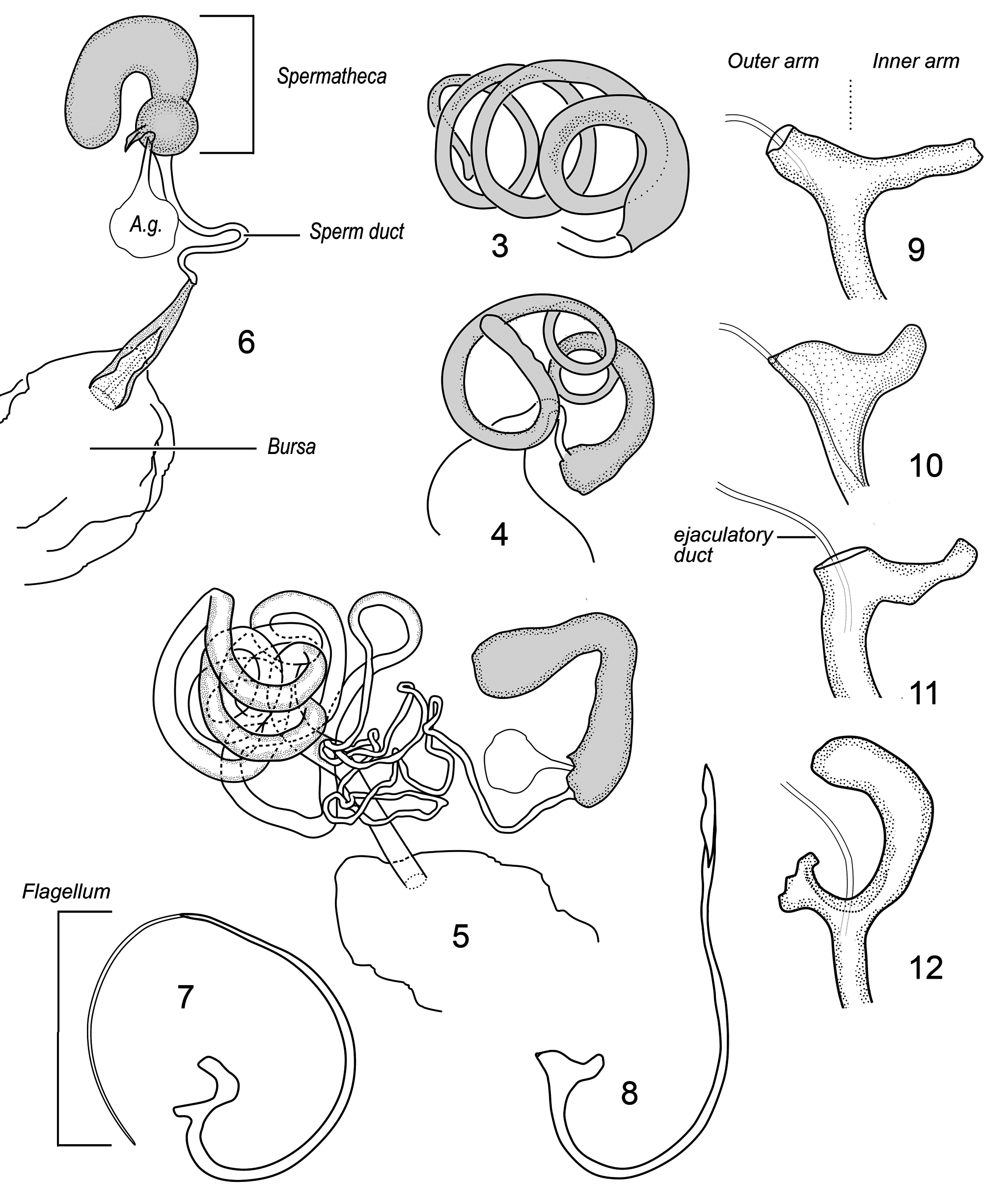
Pubescent members of the newly recircumscribed tribe Hyperaspidini (see “Remarks,” below) differ from Diomini in possessing an antenna with a fusiform club ( Figs. 2a,b View FIGURES 1–2 ) bearing a proliferation of setae in a membranous area on the inner (medially facing) surface of the terminal antennomere and smaller membranous area on the inner distal margin of the penultimate antennomere (antennae directed anteriorly); terminal maxillary palpomere in repose ( Fig. 2a View FIGURES 1–2 , left side of image) with inner edge contiguous with or partially inserted beneath anterolateral lobe of large cordate mentum, with oblique sensory surface facing dorsally and pressed against ventral surface of head, and tarsi cryptotetramerous ( Fig. 2c View FIGURES 1–2 ). Also in Hyperaspidini the abdominal postcoxal line ( Fig. 2d View FIGURES 1–2 ) does not reach the posterior margin of the ventrite and/or the outer end is recurved and directed toward or attains the anterior margin of the ventrite (see corresponding character states for Diomini ( Fig. 1 View FIGURES 1–2 ) listed in the paragraph above this one). Members of the subtribe Selvadiina ( Hyperaspidini ) can further be distinguished from Diomini based on the extremely elongate and convoluted cornu of the female spermathecal capsule ( Figs. 3–4 View FIGURES 3–12 ).
Remarks. Ślipiński (2007) diagnoses the Diomini as lacking interfacetal setae, an ocular canthus, and stylus of the coxites (=female genital plates or hemisternites) ( Table 2). Our own random sampling of Diomini exemplars in the USNM suggests that these structures are normally present, but at times difficult to see or lost through abrasion. Published digital images of Australian species also document the presence of these structures, at least in some species (e.g. Ślipiński 2007:figs. 298 & 309; Pang & Ślipiński 2010:25d, showing coxites bearing styli; Pang & Ślipiński 2009:fig. 17b, showing frontal view of specimen with eyes bearing canthi and interfacetal setae). Other character states involving the number of antennomeres in the club and the presence or absence of an infundibulum in the female genitalia are difficult to assess because no consistent method for delineating the club has been presented, and the term “infundibulum” has not been well defined nor treated consistently by various authors. When Ślipiński (2007) indicated that certain genera “probably do not belong in Diomini ” we assume from his later remarks that he intended to name the two taxa related to Selvadius — Erratodiomus and Magnodiomus —but instead mentioned Erratodiomus and Heterodiomus .
Gordon (1985) originally misinterpreted the unusual convoluted cornu of the spermathecal capsule in Selvadius ( Fig. 3 View FIGURES 3–12 ) as a continuation of the spermduct, and assumed that the spermatheca was lacking. He correctly interpreted the homologous structures in Magnodiomus and Erratodiomus ( Fig. 4 View FIGURES 3–12 ) ( Gordon 1999) but failed to consider their placement in Selvadiini , possibly due to his original misinterpretation. Gordon placed his new tribe Selvadiini in the subfamily Scymninae , but discussed similarities between Selvadius and certain hyperaspidines, even referring to them as Hyperaspidinae in his tribal diagnosis. Vandenberg (2002) indicated that Selvadiini would be better placed in the Hyperaspidinae along with Hyperaspidini and Brachiacanthini . These similarities had been previously noted by Whitehead (1967) who suggested that Selvadius may bear a closer affiliation to Hyperaspidius than to Scymnus .
In addition, Vandenberg and Perez-Gelabert (2007) noted that two South American genera, Mimoscymnus and Planorbata , originally placed in Coccidulini (Coccidulinae) ( Gordon 1994), also belong in Hyperaspidinae . Ślipiński (2007) concurred about the improper placement of Planorbata but referred to it as “a ‘Scymninae’ genus,” possibly using the latter subfamily in the broad sense of Sasaji, which would have included Hyperaspis and allied genera. Seago et al. (2011) reduced Hyperaspidinae to tribal level and placed Brachiacanthini as a synonym of Hyperaspidini . This action was followed by Gordon et al. (2014), although inconsistently within that paper, and mentioned but not employed in a subsequent work in that series ( Canepari et al. 2016). We follow Seago et al. (2011) and various other modern authors in treating the Hyperaspidini at tribal level, but recognize four distinct subtribes: Hyperaspidina, Brachiacanthina, Selvadiina stat. nov. (for Selvadius, Magnodiomu s and Erratodiomus ) and Mimoscymnina subtribe nov. (for Mimoscymnus and Planorbata ; type genus= Mimoscymnus ), as distinct lineages each defined by one or more autapomorphies of the male and female genitalia. The placement of the Selvadiina in Hyperaspidini is supported by the molecular phylogenies of Seago et al. (2011) and Robertson et al. (2015) who show Selvadius as clustering more closely with Brachiacantha and Hyperaspis than with Diomus. The placement of Mimoscymnina in Hyperaspidini is provisional, and based only on external morphological characters ( Fig. 2 View FIGURES 1–2 ) as there has not yet been a molecular study involving either Mimoscymnus or Planorbata . Members of Mimoscymnina can be readily distinguished from other members of Hyperaspidini by the elongate triangulate coxites of the female genitalia, and the male genitalia with the trabes much longer than the basal lobe and basal piece combined.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Diomini Gordon, 1999
| Vandenberg, Natalia J. & Hanson, Paul E. 2019 |
Diomini
| Gordon, R. D. 1999: 3 |