
Palaemon parvibrachium, Komai & Hanai, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5443.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:0633BE68-BB8F-4835-B658-0106DEB29D4B |
|
DOI |
https://doi.org/10.5281/zenodo.11068880 |
|
persistent identifier |
https://treatment.plazi.org/id/DF59DE4D-FFD2-F145-FF30-314F2322FEDD |
|
treatment provided by |
Plazi |
|
scientific name |
Palaemon parvibrachium |
| status |
sp. nov. |
Palaemon parvibrachium n. sp.
[New Japanese name: Amime-suji-ebi]
( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Type material. Holotype: CBM-ZC 17693, female (cl 5.8 mm; DNA voucher), Ena Bay , Miura Peninsula, 35°08'44"N, 139°39'55"E, shallow subtidal, sea grass beds of Nanozostera japonica , 1 August 2023, by dip net, coll. M. Hanai. GoogleMaps
Paratypes: CBM-ZC 17694, 1 female (cl 5.4 mm), same locality as holotype, shallow subtidal, 12 August 2022, by dip net, coll. M. Hanai GoogleMaps ; CBM-ZC 17694, 1 male (cl 4.2 mm; rostrum damaged), same data GoogleMaps ; CBM-ZC 17695, 1 male (cl 4.1 mm, rostrum damaged), same locality as holotype, shallow subtidal, 16 November 2023, by dip net, coll. M. Hanai. GoogleMaps
Diagnosis. Body integument glabrous. Rostrum 0.8–1.2 times as long as carapace, reaching or slightly overreaching distal margin of antennal scaphocerite, almost straight; dorsal margin armed with 15 teeth, including 3 postrostral, posteriormost (epigastric) tooth located at 0.2–0.25 carapace length; ventral blade well developed; no rostral crest. Carapace with branchiostegal suture originating just above base of marginal branchiostegal tooth, running straight back. Sixth pleomere without preanal tooth. Mandibular palp consisting of 3 articles. Second pereopods symmetrical, extending beyond distal margin of scaphocerite by length of fingers; carpus short, 0.8 times as long as merus, widened distally, 3.6–3.7 times as long as distal width; chela 1.8–2.0 times as long as carpus; dactylus subequal in length to palm, occlusal margin without teeth or notch. Third to fifth pereopods moderately slender; dactyli 0.3–0.4 times as long as propodi.
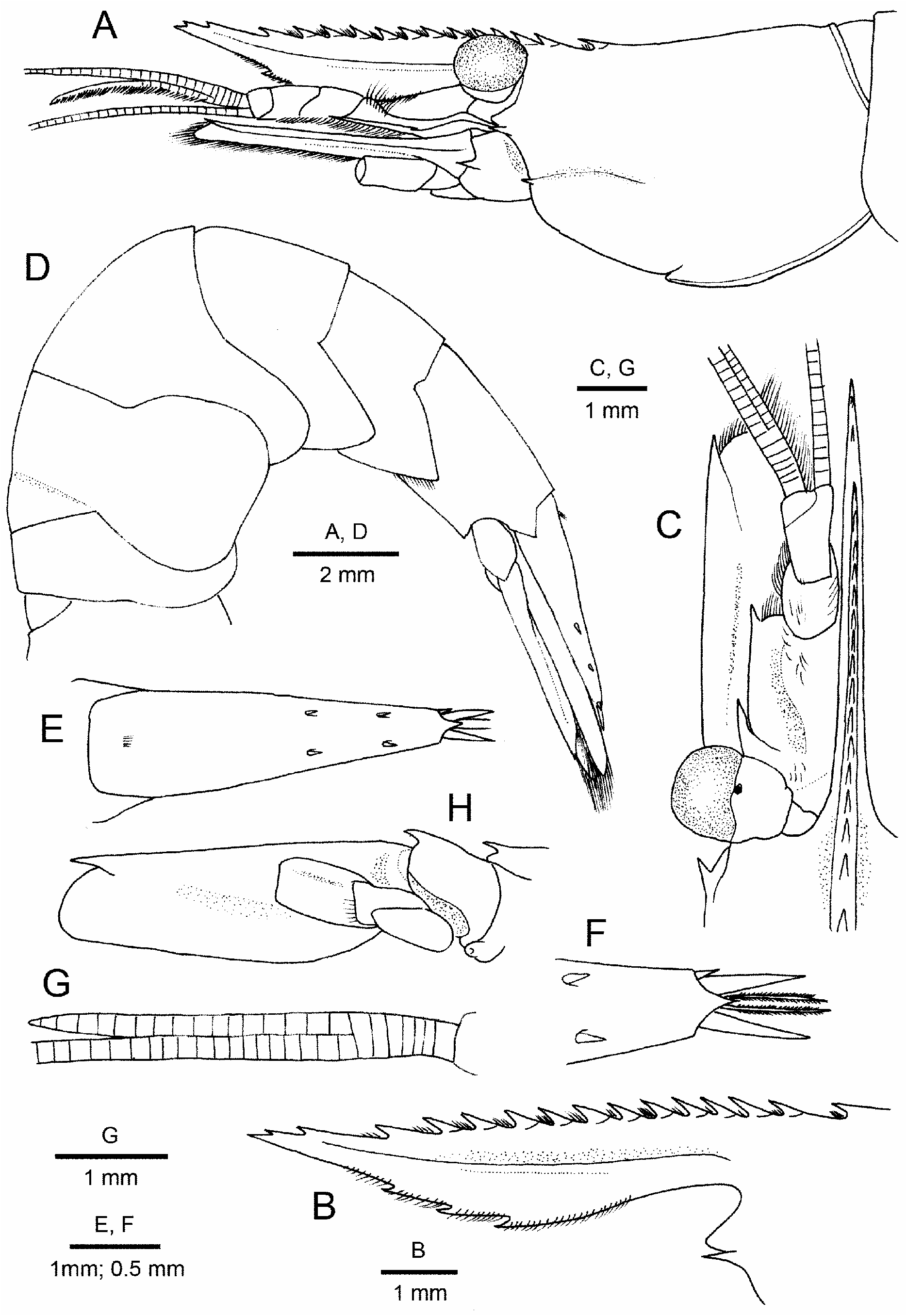
Description. Holotype. Body ( Fig. 1A, D View FIGURE 1 ) moderately slender, slightly compressed laterally; integument glabrous. Rostrum ( Figs. 1A–C View FIGURE 1 )) moderately slender, nearly straight with slightly upturned distal part, 1.2 times as long as carapace, slightly overreaching distal margin of antennal scaphocerite; dorsal margin armed along entire length with 15 teeth, including 3 postrostral and 2 subterminal teeth, posteriormost (epigastric) tooth slightly removed from preceding tooth, located at 0.2 of carapace length; proximal 13 teeth with incomplete basal suture; ventral margin gently convex, broadest at mid-length, armed with 3 teeth; spacing between remaining teeth (except for 2 subterminal teeth) roughly equal; double row of setae present in between ventral teeth, distally only a single row. Carapace ( Fig. 1A View FIGURE 1 ) dorsal profile in lateral view slightly sinuous; antennal and branchiostegal spines moderately small; branchiostegal suture originating just above base of branchiostegal spine, running straight posteriorly, terminating just in front of mid-length of carapace, same level as insertion of branchiostegal spine; suborbital lobe triangular with blunt apex, falling slightly short of antennal spine; pterygostomial angle rounded. Interocular process with upwardly directed beak, anterodorsal surface shallowly concave.
Epistome with pair of low, rounded elevations.
First to third pleonal pleura ( Fig. 1D View FIGURE 1 ) broadly rounded; fourth pleuron posteroventral margin blunt; fifth pleuron with posteroventral angle sharply pointed. Sixth pleomere approximately 1.6 times as long as fifth pleomere, 1.5 times as long as proximal height; posterolateral margin with small tooth; posterolateral process terminating acutely; preanal tooth absent. Telson ( Fig. 1D–F View FIGURE 1 ) 1.2 times as long as sixth pleomere, 3.6 times as long as proximal width, tapering distally; dorsal surface with 2 pairs of spiniform setae, anterior pair situated at 0.6 of telson length, posterior pair at 0.8 of telson length; proximal dorsal tuft of setae present; marginal setae absent; posterior margin ( Fig. 1F View FIGURE 1 ) drawn out into acute median tooth, with 1 pair of plumose setae and 2 pairs of greatly unequal spiniform setae, inner pair about 5 times longer than outer pair (left one broken off in holotype), latter not reaching tip of median tooth.
Eye ( Fig. 1A, C View FIGURE 1 ) well developed with darkly pigmented cornea; cornea slightly wider than ocular peduncle but approximately equal in length. 0.24 of carapace length; ocellar spot (nebenaugen) present at base of cornea on dorsal side.
Antennular peduncle ( Fig. 1A, C View FIGURE 1 ) falling far short of level of base of distolateral tooth of antennal scaphocerite. Basal article approximately twice as long as wide, lateral margin nearly straight; stylocerite acute, arising distinctly proximal to midlength of basal article; statocyst with statolith; distolateral tooth distinctly exceeding laminar portion; acute ventromesial tooth present. Intermediate article with noticeably produced distolateral and distomesial margins. Distal article distinctly longer than intermediate article, their combined length being about two-thirds that of basal article. Dorsal flagellum elongate, fused in 6 proximal segments, shorter ramus of 17 segments ( Fig. 1G View FIGURE 1 ), bearing numerous aesthetascs; ventral flagellum shorter than dorsal flagellum.
Antenna ( Fig. 1A, C, H View FIGURE 1 ) with scaphocerite moderately broad, 2.9 times as long as broad. 0.9 times as long as carapace; outer margin straight, terminating in tooth, falling short of rounded distal margin of lamella. Basicerite with small lateral tooth. Carpocerite falling short of midlength of scaphocerite. Flagellum longer than body.
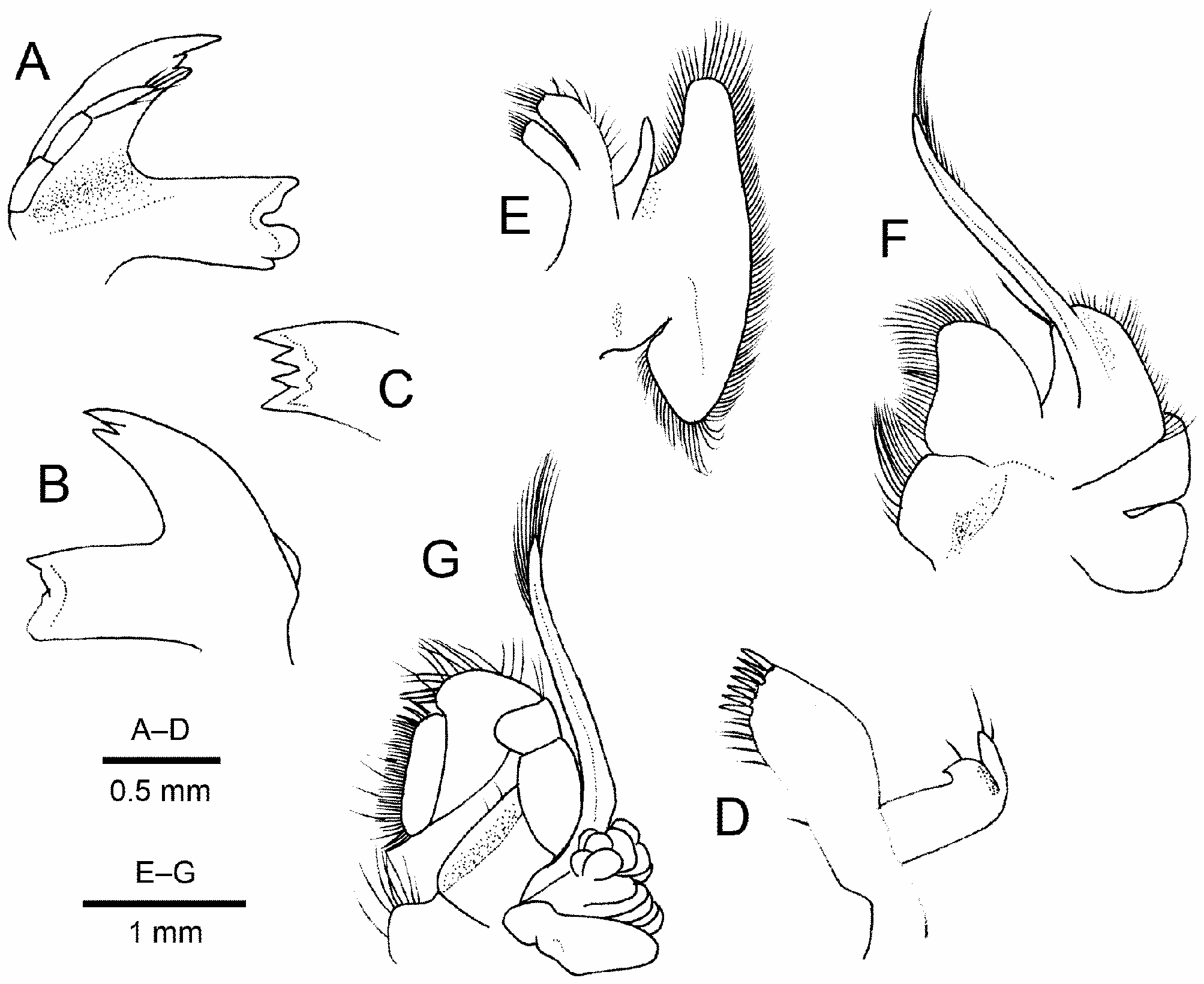
Mouthparts quite typical for genus. Mandible ( Fig. 2A–C View FIGURE 2 ) with palp consisting of 3 articles, third article longest, with some distal setae; incisor process with 4 distal teeth; molar process distally 4-toothed. Maxillule ( Fig. 2D View FIGURE 2 ) suboval basial endite, bearing double row of spiniform setae on truncate distal margin; endopod bilobed distally, inner lobe recurved, outer bluntly pointed, with 2 short setae distally. Maxilla ( Fig. 2E View FIGURE 2 ) with poorly developed coxal endite; basial endite deeply bilobed; endopod feebly curved mesially, terminating in subacute tip; scaphognathite moderately broad. First maxilliped ( Fig. 2F View FIGURE 2 ) with coxal and basial endites moderately broad; endopod somewhat curved inward, reaching as far as distal margin of basial endite, tapering to subacute tip; caridean lobe moderately broad, with long flagellum; epipod deeply bilobed. Second maxilliped ( Fig. 2G View FIGURE 2 ) with carpus rounded at inner angle; epipod subovate, with well-developed podobranch.
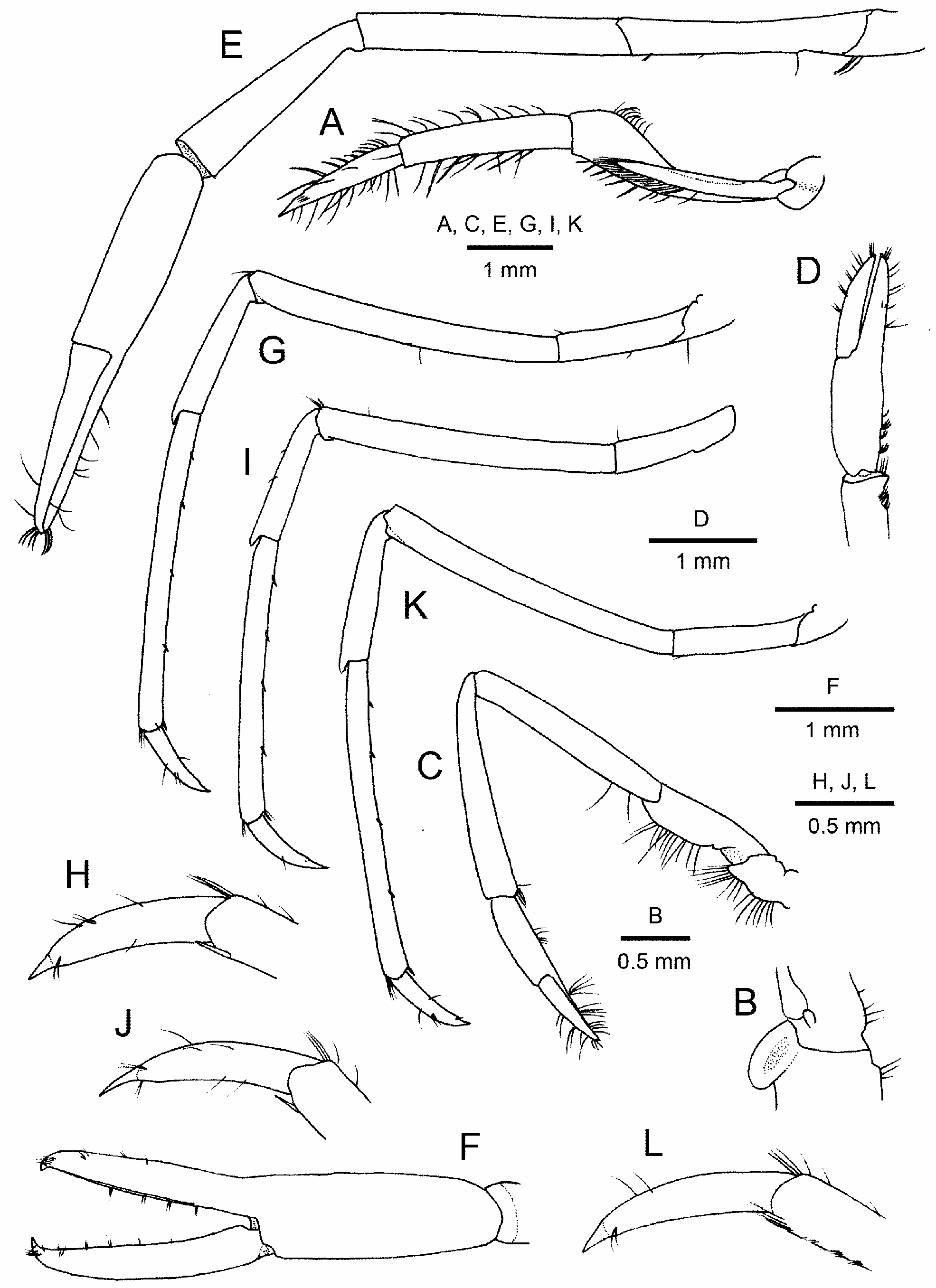
Third maxilliped ( Fig. 3A, B View FIGURE 3 ) reaching to middle of third segment of antennular peduncle, generally compressed laterally. Coxa with broadly rounded lateral plate. Antepenultimate article strongly flattened proximally, strongly thickened distally. Penultimate article (= carpus) subcylindrical, slightly shorter than antepenultimate article, with sparse stiff setae on extensor and flexor margins. Ultimate article tapering to acute tip, 0.8 times as long as penultimate article, with transverse tracts of stiff setae on mesial face.
Well-developed pleurobranchs present above bases of first to fifth pereopods.
First pereopod ( Fig. 3C, D View FIGURE 3 ) falling slightly short of distal margin of scaphocerite. Basis distinctly shorter than ischium. Ischium broader than merus, with distinct notch on ventral margin proximally; ventral margin distal to proximal notch gently convex. Merus 1.7 length as long as ischium. Carpus as long as merus, slightly widened distally. Chela 0.80 length of carpus; fingers subequal in length to palm, with tufts of setae; occlusal margins straight, unarmed. Carpal-propodal brush, consisting of grooming apparatus, well developed.
Second pereopods ( Fig. 3E, F View FIGURE 3 ) symmetrical, extending beyond distal margin of scaphocerite by length of fingers. Merus subequal in length to ischium, 6.9 times as long as wide. Carpus short, 0.8 times as long as merus, widened distally, 3,7 times as long as distal width. Chela twice as long as carpus; palm 2.8 times as long as wide; fingers terminating in hooked, acute tips; fixed finger occlusal margin without tooth or notch; dactylus subequal in length to palm, occlusal margin without teeth or notch.
Ambulatory pereopods generally similar, moderately slender, increasing in length from third to fifth. Third pereopod ( Fig. 3G, H View FIGURE 3 ) just reaching distal margin of scaphocerite by tip of dactylus. Merus 2.3 times as long as ischium. Carpus half-length of merus. Propodus twice length of carpus, subequal in length to merus, flexor margin armed with 4 widely spaced, single spiniform setae and a distal pair. Dactylus ( Fig. 3H View FIGURE 3 ) tapering, distinctly narrowing at base of unguis, slightly curved, 0.3 times as long as propodus, 5 times as long as proximal width; unguis slender.
Fourth pereopod ( Fig. 3I, J View FIGURE 3 ) similar to third pereopod, overreaching distal margin of scaphocerite by about half length of dactylus.
Fifth pereopod ( Fig. 3K, L View FIGURE 3 ) overreaching distal margin of scaphocerite by half length of dactylus. Merus 2.2 times as long as ischium. Carpus half-length of merus. Propodus 2.1 times as long as carpus, flexor margin armed with 4 widely spaced spiniform setae, including one on mesiodistal margin; propodal grooming brush weakly developed, comprising 5 rows of 2–5 serrulate setae, extending for about distal 0.2 length of propodus. Dactylus tapering, distinctly narrowed at base of unguis, gently curved, 0.3 times as long as propodus, 4.9 times as long as proximal width.
Uropods ( Fig. 1D View FIGURE 1 ) overreaching telson by 0.2 length of endopod. Exopod subequal in length to endopod, diaeresis reaching beyond midline; posterolateral spiniform seta longer than posterolateral tooth.
Paratypes. Generally similar to holotype. Rostrum 0.8 times as long as carapace, with 15 dorsal teeth (female paratype) including 3 postrostral, posteriormost tooth located at 0.24–0.25 of carapace length; ventral margin with 3 teeth (female paratype). Second pereopod with carpus short, 0.8 times as long as merus, widened distally, 3.6 times as long as distal width; chela 1.8 times as long as carpus; palm 2.5 times as long as wide; dactylus subequal in length to palm. Dactyli of third to fifth pereopods 0.3–0.4 times as long as propodi.
Colouration in life ( Figs. 4–5 View FIGURE 4 View FIGURE 5 ). Generally transparent with pattern of brown lines and dots. Rostrum with line of brown dots along lateral carina. Carapace with conspicuous lines of brown dots obliquely across branchiostegite from postorbital region, antennal tooth and branchiostegal tooth; cardiac region with short lines of brown dots. Pleon with transverse lines and dots on each pleomeres, transverse line along posterior margin of third pleomere distinct. Cornea grey. Antennular and antennal flagella brown. Second pereopod with tinge of light blue and brown dots, base of fingers yellowish. First and third to fifth pereopods translucent, with brown dots on each article.
Etymology. From the combination of the Latin parvus (= small, short) and brachium (= forearm), referring to the characteristically short carpus of the second pereopod of this new species. Used as a noun in apposition.
Distribution. Presently known only from the type locality, Ena Bay, Miura Peninsula; intertidal to shallow subtidal.
Habitat. All specimens of the new species were collected from seaweed beds of Nanozostera japonica at low tide ( Fig. 6 View FIGURE 6 ). Two congeneric species, P. macrodactylus Rathbun, 1902 and P. ortmanni Rathbun, 1902 , were also encountered sympatrically at the collection site.
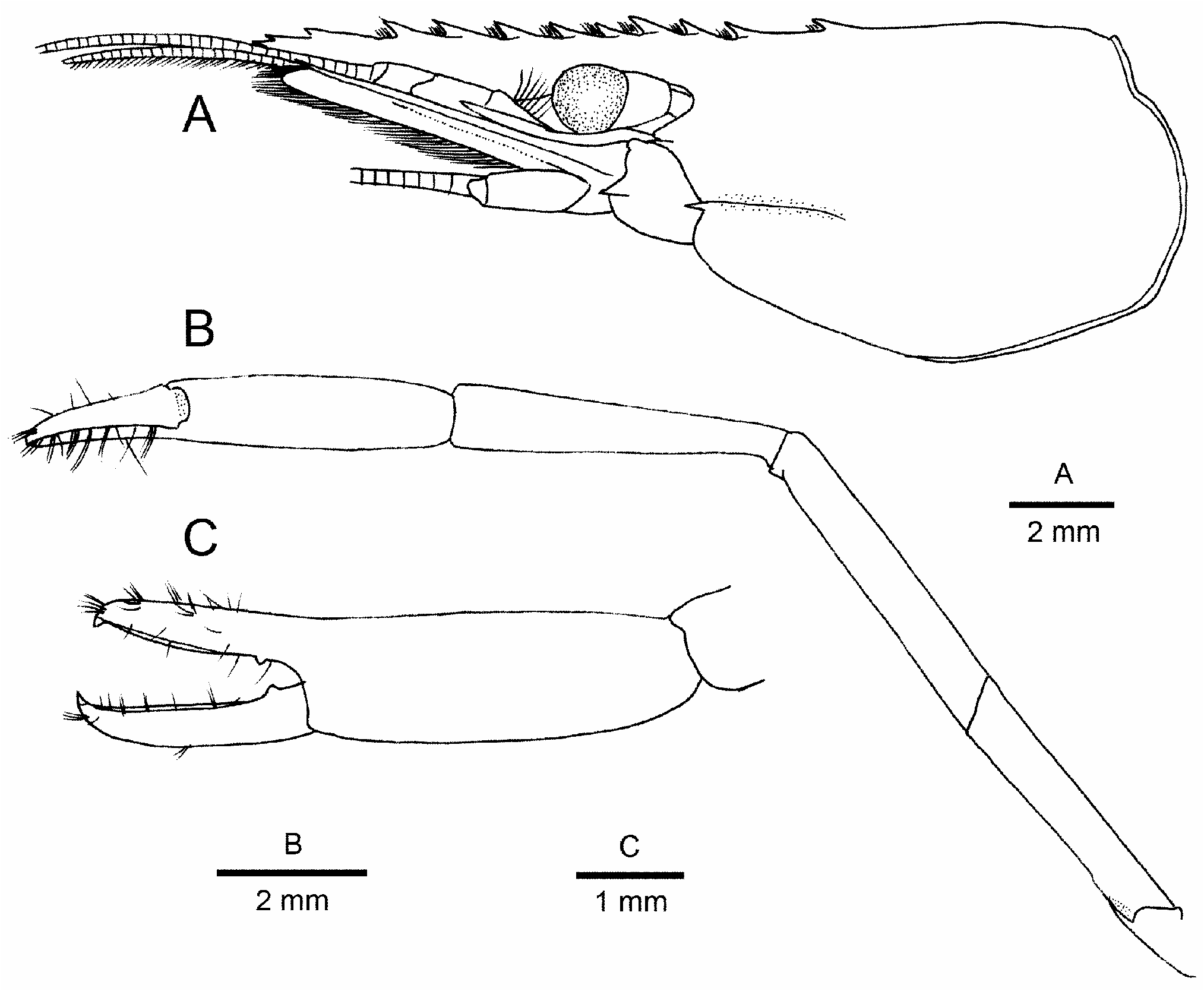
Remarks. Morphologically, the present new species appears most similar to Palaemon serrifer , having been recorded widely from the Indo-West Pacific region (e.g., Holthuis 1950a; Anker & De Grave 2016). Stimpson’s (1860) original description of P. serrifer (as Leander ) was based on specimens from Hong Kong and “Oushima” (= Amami-ohshima Island, Ryukyu Islands, Japan). Bruce (1986) redescribed P. serrifer with topotypic specimens from Hong Kong. Previous phylogenetic studies ( Ashelby et al. 2012; Carvalho et al. 2017) suggest that more than one species are mixed up under material identified with P. serrifer , and in fact there are two nominal taxa ( Leander fagei Yu, 1930 and Leander serrifer var. longidactylus Yu, 1930 ), which have been synonymized under P. serrifer by Holthuis (1950a). A reappraisal of the identity of P. serrifer will be necessary. In this study, we examined Japanese specimens agreeing with the descriptions of P. serrifer by Kubo (1942), Holthuis (1950a), Hayashi (2000a), Li et al. (2007) for comparison (see above “Material and methods” section). COI and/or 16S rRNA gene sequences have been generated from four specimens, including one topotypic specimen from Amami-ohshima Island, Ryukyu Islands ( Table 1 View TABLE 1 ). Of them, 16S rRNA sequences match well JQ042295 registered in GenBank (>99.4% identity; see Table 2 View TABLE 2 , voucher specimen from the Seto Inland Sea, Japan). Other sequences of questionable identity were not included in the present genetic analysis (JN674348, KC515048 and MN993996 for the 16S rRNA gene; JX502985 and ON753717 for the COI gene).
Shared diagnostic characters between P. parvibrachium n. sp. and P. serrifer include: rostrum not particularly elongate, slightly overreaching distal margin of scaphocerite at most, nearly straight; branchiostegal suture of carapace originating just above base of antennal spine; carpus of second pereopod shorter than chela; dactyli of third and fourth pereopods about 0.3 times as long as propodi. Nevertheless, the new species is readily distinguished from P. serrifer by the following particulars: (1) the epigastric tooth (the posteriormost tooth of the dorsal rostral series) is more anteriorly situated in P. parvibrachium n. sp. than in P. serrifer (0.2–0.25 length of the carapace versus 0.3 length) (compare Fig. 1A View FIGURE 1 vs. Fig. 7A View FIGURE 7 ); (2) the carpus of the second pereopod is proportionally shorter (about half length of chela versus 0.8 length) (compare Fig. 3E View FIGURE 3 vs. Fig. 7B View FIGURE 7 ); and (3) the dactylus of the second pereopod is proportionally longer (subequal in length to palm versus distinctly shorter than palm) (compare Fig. 3F View FIGURE 3 vs. Fig. 7 View FIGURE 7 ). The live colouration is also different between the new species and P. serrifer . In the new species, there are more numerous lines on the carapace and pleon; those lines are brown in P. parvibrachium n. sp., whereas dark brown to black in P. serrifer (compare Figs. 4–5 View FIGURE 4 View FIGURE 5 vs. Fig. 8 View FIGURE 8 ). Genetically, the new species is substantially distant from P. serrifer : genetic divergences between the two taxa are 20% in the COI gene and 6.6–9.1% in the 16S rRNA gene.
There were no highly matching sequences among Palaemon parvibrachium n. sp. and the congeners registered in GenBank, although sequence data of the COI and/or 16S rRNA genes are available for 71 congeners in total ( Ashelby et al. 2012; Calvalho et al. 2017; Giraldes et al. 2024). The most similar sequences were of Palaemon gravieri ( Yu, 1930) , P. macrodactylus ( Rathbun, 1902) and P. tenuidactylus (Liu, Liang & Yan, 1990a) for the COI gene (the genetic identity 88%), and P. gravieri , P. macrodactylus , P. ogasawaraensis Kato & Takeda, 1980 and P. tenuidactylus for the 16S rRNA gene (the genetic identity <97%). None of these species show a close morphological similarity to P. parvibrachium n. sp.
No genetic data are available for the following congeners: P. africanus ( Balss, 1916) , P. audouini Heller, 1861 , P. colossus Tzomos & Koukouras, 2015 , P. cummingi ( Chace, 1954) , P. curvirostris Nguyên, 1992 , P. gladiator Holthuis, 1950b , P. hainanensis ( Liang, 2000) , P. hiltoni ( Schmitt, 1921) , P. hobbsi ( Strenth, 1994) , P. maculatus ( Thallwitz, 1892) , P. mani ( Sollaud, 1914) , P. mesogenitor ( Sollaud, 1912) , P. miyadii Kubo, 1938 , P. peruanus Holthuis, 1950b , P. powelli Ashelby & De Grave, 2009 , P. sewelli Kemp, 1925 and P. yamashitai Fujino & Miyake, 1970 . None of these species exhibit close morphological similarity to the present new species (see respective original descriptions; Ashelby & De Grave 2009; De Grave & Ashelby 2020).
In spite of the efforts by the junior author and his colleagues, only four specimens of P. parvibrachium n. sp. have been collected during two-year (2022–2023) field surveys in Ena Bay and nearby areas. The senior author has tried to find specimens of the new species in the decapod collections housed in CBM, but this was unsuccessful. At present, the true geographical distribution of the new species remains somewhat unknown, in spite of its occurrence in easily accessible coastal area.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |