
Gastrotheca litonedis Duellman & Hillis 1987
|
publication ID |
https://doi.org/10.11646/zootaxa.4562.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:DDB73CA2-F300-4C72-B936-A6685ED775AE |
|
DOI |
https://doi.org/10.5281/zenodo.5941951 |
|
persistent identifier |
https://treatment.plazi.org/id/DF65A94B-0B02-D63E-FF21-73C2F134F837 |
|
treatment provided by |
Plazi |
|
scientific name |
Gastrotheca litonedis Duellman & Hillis 1987 |
| status |
|
Gastrotheca litonedis Duellman & Hillis 1987 View in CoL
Holotype. KU 202690 ( Figs. 10A View FIGURE 10 , 25A View FIGURE 25 ), an adult female, from 10 km (by road) northeast of Girón , 2750 m ( 03° 07' 11.8" S, 79° 06' 28.4'' W), Azuay Province, Ecuador, obtained on 7 March 1984 by William E. Duellman. GoogleMaps
Referred specimens. (Total 28: 13 males, 15 females). Ecuador: Azuay: CJ 386 ¯9 (females), 401¯4 (females, male), 1404¯8 (males) , KU 335388–9 (male, female), San Fernando , Laguna de Busa, 2834 m ( 03° 09' 15.8" S, 79° 15' 49.03" W), collected on 9 June 2011 by Elicio E. Tapia, Sofía Carvajal-Endara and Henry Grefa GoogleMaps ; QCAZ 42734 View Materials (female), San Fernando , Laguna de Busa, 2834 m ( 03° 09' 15.8" S, 79° 15' 49.03" W), collected on 27 October 2007 by Sofía Carvajal-Endara, Amaranta Carvajal-Campos and Andrea Carvajal-Endara GoogleMaps ; QCAZ 42855 View Materials (female), 42857¯9 (females), 42861 (female), 42866 (male), 42871 (male), San Fernando , Laguna de Busa, 2834 m ( 03° 09' 15.8" S, 79° 15' 49.03" W), collected on 18–20 August 2008 by Sofía Carvajal-Endara GoogleMaps ; QCAZ 49973 View Materials ¯4 (female, male), 49976¯8 (males), San Gerardo , 2854 m ( 03° 08' 00.17" S, 79° 11' 36.78" W), collected on 1 December 2010 by Sofía Carvajal-Endara GoogleMaps .
Diagnosis. Included in the genus Gastrotheca by having a closed brood pouch on dorsum of female. A moderately large species ( 53.5– 62.4 mm SVL in females, n = 22; 48.9–57.4 mm SVL in males, n = 13) with tibia length 40¯54% SVL, slightly longer than foot; (2) interorbital distance about 1.5 times width of upper eyelid; (3) skin on dorsum smooth, not co-ossified with skull, lacking transverse ridges; (4) supraciliary processes absent; (5) heel lacking calcar or tubercle; (6) tympanic annulus distinct, smooth; (7) Fingers I and II about equal in length, width of discs notably wider than digits; (8) fingers unwebbed; (9) webbing between external toes extending to the penultimate subarticular tubercle on Toe IV, to the distal subarticular tubercle on Toe V; (10) in life, dorsum green, brown, or tan with or without dark paravertebral marks or dark flecks; (11) head markings consisting of a pale cream labial stripe and narrow dark brown canthal stripe; (12) pale creamy white dorsolateral stripe present; (13) flanks, anterior surfaces of thighs and groin pale brown or tan slightly molted in some, posterior surfaces of thighs dark brown–gray with pale cream warts in some; (14) venter pale brownish gray; (15) brood pouch single, dorsal.
Gastrotheca litonedis most closely resembles five other species in southern Ecuador— G. cuencana , G. plumbea , G. pseustes , G. turnerorum , and G. elicioi . Gastrotheca litonedis differs from all of them by having a pale brownish gray venter; whereas, the venter is uniform creamy white in G. cuencana , cream in G. plumbea , cream with dark flecks or spots in G. pseustes and G. elicioi , and bronze or dark brown in G. turnerorum . Gastrotheca litonedis , like G. plumbea , has smooth skin on the dorsum, whereas it is finely granular in G. elicioi and G. cuencana , weakly areolate to smooth in G. pseustes and areolate in G. turnerorum . A pale supracloacal stripe is present in G. litonedis , G. pseustes , and G. turnerorum , but it is absent in G. cuencana and G. plumbea . Gastrotheca pseustes differs further from G. litonedis by having a complex call structure (long and short pulsed-notes), which is simple in G. litonedis (only short pulsed-notes). The sister species of G. litonedis is the parapatric G. cuencana . Distinctive morphological and coloration features (compare Figs. 26–27 View FIGURE 26 View FIGURE 27 , vs 2–4) are (character states for G. cuencana in parentheses): Gastrotheca litonedis is larger than G. cuencana (SVL student’s t-test: t = -6.725, df = 99, p <0.001), has a bluntly rounded snout in profile (rounded snout inclined anteroventrally in profile); a narrow dark brown canthal stripe is present (absent or inconspicuous), a row of barely elevated dorsolateral warts (conspicuous, elevated), green, tan or brown dorsal surfaces of fingers (fingers cream dorsally), posterior surfaces of thighs dark brown (translucent cream with pale tinge blue or green). The call of G. litonedis has a higher note rate, longer note duration, shorter inter-note interval, higher dominant frequency, and higher 90% bandwidth frequency compared with G. cuencana . Finally, these two species also differ by having a genetic distance of 1.2 % (in a DNA dataset of 438 bp, 16S gene).
Gastrotheca litonedis is syntopic with G. lojana and G. pseustes 2, from which it differs notably by the texture of skin on dorsum, ventral coloration, and call structure. The skin on the dorsum is smooth in G. litonedis , whereas it is finely granular in G. lojana and coarsely granular in G. pseustes 2; furthermore, both species have cream venters with dark flecks, spots, or mottling, and a complex call structure with long and short pulsed-notes ( G. lojana call in Fig. 17 View FIGURE 17 H–N).
Variation. Morphometric variation of 22 females and 13 males (from Duellman 2015) is summarized in Table 3. Females are larger than males (58.0± 2.4 mm; 53.8± 2.8 mm). The skin on the dorsum is mostly smooth in most specimens; however, in some specimens it is weakly areolate. All adults have a moderate supratympanic fold, which usually extends from the upper part of the tympanum to a point above the insertion of the forelimb. Some individuals have several small rounded tubercles anterolateral to the tympanum. Each dentigerous vomerine process has 2–10 teeth (5.4±1.6, n = 34).
Color variation in preservative. In most specimens the dorsum is bluish gray without marks; in a few specimens faint paravertebral marks are present. The narrow canthal stripe, flanks, and posterior surfaces of thighs are dark brown. A creamy white supralabial stripe from the posterior margin of the lip to the insertion of the forelimb is prominent in all specimens. Creamy white dorsolateral, supracloacal and heel stripes are present. On the posterior part of the flanks the coloration varies from small white flecks to mottling on a dark brown ground color. The anterior surfaces of the thighs are usually gray with black flecks or spots. The ventral surfaces are uniform dull gray in most specimens; some have a pale gray venter with evenly distributed dark flecks and spots, and in CJ 1412 the venter is pale gray with black markings. Some specimens have a faint tinge of pale blue on the ventral surfaces of the shanks. Males have a cream nuptial pad on the medial surface of the thumb.
Color variation in life. ( Figs. 26 View FIGURE 26 , 27 View FIGURE 27 ). The dorsum varies from uniform green (QCAZ 42858), to brown (CJ 404); in some individuals dark paravertebral marks and dark brown flecks are present (QCAZ 49977). The ventral surfaces are pale gray in most specimens; some have evenly distributed dark flecks and spots, and some have a faint bluish tinge on the ventral surfaces (QCAZ 49977). A narrow dark brown canthal stripe is present; a cream supralabial stripe extends from the posterior margin of the lip to the insertion of the forelimb. The tympanum is brown, tan, or olive green. The iris is reddish bronze with dense black reticulations. The posterior surfaces of the thighs are dark brown or cream with orange flecks. A cream dorsolateral stripe is evident in most individuals. The flanks, groin, and anterior surface of the thighs usually are bronze-brown or tan with cream or dark flecks and spots. In some individuals a pink-orange tinge is present in the groin ( Fig. 27 View FIGURE 27 ). The supracloacal and heel stripes are cream or tan, and the ventral surfaces of the shanks have a faint pale blue tinge.
Tadpoles. Tadpoles belong to Type IV tadpole of Orton (1953), and the exotroph, benthic ecomorphological guild. All measurements are expressed in millimeters. The following description is based on a specimen in Stage 37 (CJ 6558), from a series of 79 tadpoles (CJ 6557) born and reared in laboratory conditions, from a mother collected when gravid in the field (CJ 6018) at Laguna de Busa, 2780 m, Azuay Province, Ecuador, by Manuel A. Morales-Mite on 0 6 January 2016 ( Fig. 5A View FIGURE 5 ).
Total length 52.8; body length 19.4 (37% of total length). Body ovoid in dorsal and lateral views, slightly depressed; throat slightly concave in lateral profile, sloping from tip of snout to belly; body width at the level of spiracle 12.5, and height at same position 10.3, head width at level of eyes 10.0. Lateral line system present but barely visible, supraorbital and infraorbital lines not evident at level of snout, infraorbital line present at level of the eye, touching the inferior portion of the orbit, and making contact with supraorbital line immediately behind the eye. Postorbital line represented by three stitches arranged in line and forming a 45-degree angle with the body plane. Angular line, anterior oral line and loreal lines not visible; dorsal body and middle body lines not visible. Nostril medium sized (in proportion to body length), ovoid, protruding, having a fleshy annulus, its opening directed anterolaterally. Snout–nostril distance 4.2; internarial distance 2.8. Eyes dorsally positioned; eye length 1.9, eye width 1.8; interorbital distance 5.2. Spiracle sinistral, located at midbody level, spiracular opening oriented posteriorly; distance from tip of snout to spiracular opening 12.9; end of spiracular tube rounded, attached to body wall, inner wall of spiracular tube not evident; spiracle length 2.7, spiracle transverse width 3.5. Vent tube dextral, opening oriented posteriorly, tube length 2.5, tube transverse width 2.2. Tail length 33.3; caudal musculature slender, narrowing gradually until tail terminus; caudal muscle height 4.1, width 3.6; caudal fins well developed and proportional, dorsal fin arising abruptly near tail-body junction; dorsal fin height 4.1, ventral fin height 3.7; maximum tail height 9.2; tail terminus rounded, caudal musculature not reaching fin terminus.
Oral disc small, ventral, located near tip of snout, not protruding laterally beyond body; transverse width 5.8. It is surrounded by a uniserial row of marginal papillae, interrupted medially on upper lip. Lower lip papillae alternate in orientation, giving appearance of two rows; upper lip with 23 papillae on right side and 23 papillae on left side; lower lip bearing 57 marginal papillae. Upper jaw sheath medium-sized, forming a smooth arch and finely serrated, transverse width 2.9 (50% of width of oral width) and height 0.3. Lower jaw sheath V- shaped, open and finely serrated, width 2.6, and height 0.7. Labial tooth row formula 2/3(1); tooth rows lengths: A1: 4.5, A2: 4.4, P1 right row 1.65, P1 left row 2.0, P1 gap 0.2, P2: 4.1, P3: 3.95. (See also Fig. 6A View FIGURE 6 of CJ 5292, Stage 35).
Color in life. Based on a specimen in Stage 34 from a series (CJ 1947) obtained from San Fernando, Laguna de Busa, Azuay Province, 2834 m, by Elicio E. Tapia, Sofía Carvajal-Endara, and Henry Grefa on 9 June 2011 ( Fig. 6A View FIGURE 6 ). In dorsal view, body olive-gray, head and tail suffused with pink and cream. Flanks cream anteriorly to olivegray posteriorly. In ventral view, cream anteriorly in gular region; followed posteriorly by red gills visible troughout the throat, and olive-gray guts. Caudal musculature pink proximally, gradually becoming gray posteriorly; dorsal and ventral fins translucent, with olive stippling distributed regularly along length of tail except distally, where stippling diminishes. Spiracle and oral apparatus nearly translucent. Iris gold.
Variation. Variation of 28 meristic characters of tadpoles in Stages 33–40 (CJ 5447, 5449, 5464–67, 6557, 6558) are shown in Table 8. Total length varies between 31.4 (Stage 33) and 61.9 (Stage 40) and tail length proportion varies from 58% to 69% until Stage 40. Number of marginal papillae varies among specimens and Gosner stages; variation in number of ventral papillae at lower lip is high (39–66).
We documented changes in coloration during ontogenetic development of CJ 1947 (Fig. 28A–C). At Stage 39, the dorsum and flanks are olive-gray with a diffuse pattern of gray paravertebral marks. By Stage 41, dorsum of body and limbs is green with well-defined dark green paravertebral marks; a cream dorsolateral stripe is bordered below by a fine black line; a creamy white stripe is present on face where upper lip will be formed. The flanks are greenish gray. At Stage 46, the dorsum is nearly uniform green; the paraverterbal marks are barely evident. The flanks are brown, and the iris is red. CJ 6813 is similar to the above described but has scattered cream flecks on a green dorsum.
Comparisons. Tadpoles of Gastrotheca litonedis may occur in sympatry with those of G. pseustes 2 and G. lojana . Gastrotheca litonedis differs from them by having less pigmentation on dorsum and venter (compare in Fig. 5 View FIGURE 5 ). It further differs from G. lojana by having a lowest tail dorsal fin.and from G. pseustes by lacking a reticulated pattern on the flanks.
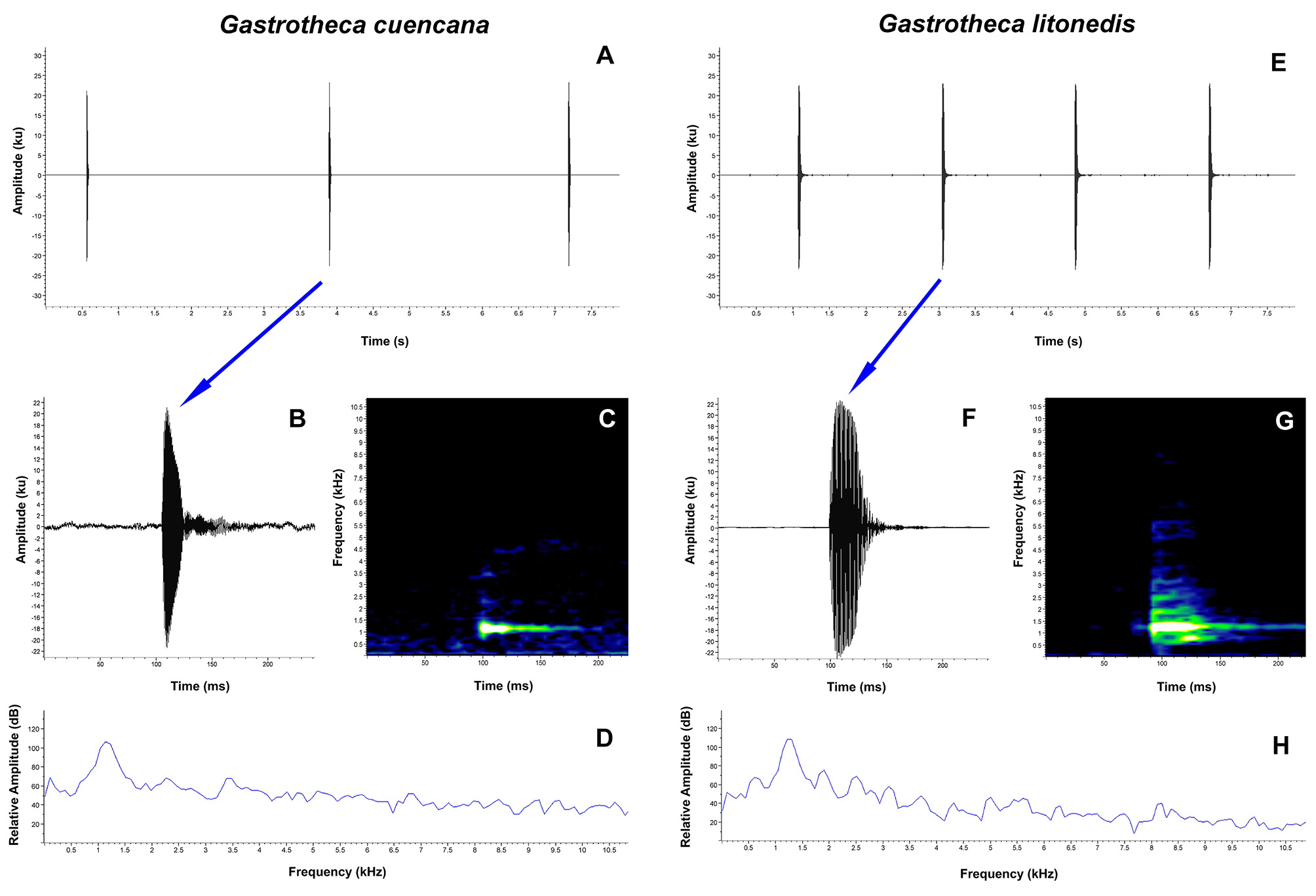
Vocalization. A total of nine individuals of Gastrotheca litonedis were recorded from two locations (6 individuals from San Gerardo and 3 from San Fernando, Laguna de Busa) in Azuay Province (Appendix III). Descriptive statistics of the acoustic variables are provided in Table 5. The advertisement call of G. litonedis is a simple call, composed of a series of short, single-pulsed notes, emitted at regular intervals ( Fig. 8 View FIGURE 8 E–H). In our recordings, the calls have between 1 and 21 notes per call. The notes have a mean duration of 0.075 s (SD = 0.025) and a mean inter-note interval of 1.768 s (SD = 0.286). The mean dominant frequency of the call is 1224.0 Hz (SD = 66.415), with mean 90% bandwidth of 1114.7–1308.1 Hz. The fundamental frequency and harmonics are not clearly recognizable.
Comparisons. The advertisement call of Gastrotheca litonedis is similar to that of G. cuencana ; all the other species of Gastrotheca species in the southern Ecuadorian Andes have complex calls. However, the call of G. litonedis has a higher note rate, longer note duration, shorter inter-note interval, higher dominant frequency, and higher 90% bandwidth frequency compared with G. cuencana ( Table 5).
Distribution and ecology. Gastrotheca litonedis is known from three localities (ca. Girón, San Fernando (Laguna de Busa), San Gerardo) on the eastern slope of the Cordillera Occidental in Azuay Province ( Fig. 9 View FIGURE 9 ). Its elevational range is 2750–2854 m in an area of extent of occurrence of about 10.2 km 2. This nocturnal, semiarboreal species inhabits forest and disturbed areas in the Evergreen Montane Forest of Cordillera Occidental of the Andes (Ministerio de Ambiente del Ecuador 2012), where the average annual rainfall is 611–658 mm and the average annual temperature is 12.2–12.5 °C ( Fick & Hijmans 2017). Most of the specimens collected at Laguna de Busa were found among totora reeds ( Schoenoplectus californicus ) at the border of the lagoon. Males were calling from totora reeds at approximately 1–2 m above ground. Females were perched on totora reeds approximately 40 cm above ground and along small streams that were close to the lagoon. Some males also were calling from branches of shrubs around the lagoon approximately 2.5 m above ground. During the day a male and a female were basking on leaves of calla lilies ( Zantedeschia aethiopica ), which were present along irrigation ditches, bordering pastures. At this locality, Gastrotheca litonedis is syntopic with G. pseustes ( sensu lato) and G. lojana ; all of them have a biphasic mode of development, in which brooding females release tadpoles in ponds, where they complete their development ( Figs. 10–11 View FIGURE 10 View FIGURE 11 ). In all the visits made to this locality ( October 2007 and December 2007, August 2008, June 2011), gravid females were found, as well as tadpoles in small streams, temporary ponds, and at the edge of the lagoon. One of the gravid females collected on 9 June 2011 was transported to the laboratory, where it deposited 96 tadpoles. At San Gerardo, most individuals were found on the border of an irrigation ditch in a pasture. Males were calling from the grass at approximately 50 cm above ground. At this locality, G. pseustes ( sensu lato) also was found. A brooding female of G. litonedis , showing heliophilic behavior during most of the incubation time, under captive conditions in an outdoor enclosure, is depicted in Figure 10C View FIGURE 10 .
Conservation status. IUCN categorizes Gastrotheca litonedis as Endangered ( Coloma et al. 2004). However, the current assessment was based on information combining the two species described in this study ( G. litonedis and G. cuencana ). Thus, we re-categorize this species as Critically Endangered, according with IUCN criteria and sub-criteria B1ab(i,ii,iii,iv). Currently, the known area of occurrence of G. litonedis is extremely small ( 10.2 km 2), and its area of occupancy is even smaller; although we suspect that its distribution might increase as further searches are conducted. Its populations are in heavily human populated areas. For example, the surroundings of Laguna de Busa have been deforested and modified for human activities ( Fig. 11D View FIGURE 11 ). The lake contains two introduced, predatory fish (trout and carp) and its surroundings are greatly altered by the introduction of exotic eucalyptus and conifers. Additional threats at this site are unregulated tourisms activities.
Comments. In the original description of Gastrotheca litonedis, Duellman and Hillis (1987) included data on nine males and 15 females. As noted by Duellman (2015), of these specimens, only the holotype, KU 202690, is G. litonedis . The other specimens used by Duellman & Hillis (1987) are identified herein as G. cuencana . In addition, the photograph published as the holotype of G. litonedis by Duellman and Hillis (1987:156: Fig. 9 View FIGURE 9 ) is not the holotype. The frog in the photograph is actually G. lojana (KU 148794, from 2 km SSW of Saraguro, Loja Province, Ecuador). Also, the frog QCAZ 2692 depicted in Duellman (2015: Fig. 11.12B) is G. lojana (not G. litonedis ). Genetic sequences generated in this study from tissues of the holotype differ from the sequences of other specimens generated and published by Wiens et al. (2007) (Genbank numbers: DQ679395 View Materials , DQ679355 View Materials , DQ679323 View Materials , DQ679287 View Materials ), which are more similar to sequences of G. cuencana .









| KU |
Biodiversity Institute, University of Kansas |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |