
Gastrotheca cuencana, Carvajal-Endara & Coloma & Morales-Mite & Guayasamin & Székely & Duellman, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4562.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:DDB73CA2-F300-4C72-B936-A6685ED775AE |
|
DOI |
https://doi.org/10.5281/zenodo.5941943 |
|
persistent identifier |
https://treatment.plazi.org/id/DF65A94B-0B2E-D65D-FF21-75D1F7F0FE23 |
|
treatment provided by |
Plazi |
|
scientific name |
Gastrotheca cuencana |
| status |
sp. nov. |
Gastrotheca cuencana View in CoL sp. nov.
urn:lsid:zoobank.org:act:
Holotype. CJ 1391 ( Fig. 2 View FIGURE 2 ), an adult male, from the city of Cuenca , 2579 m ( 02° 53' 57.91" S, 79° 01' 52.79" W), Azuay Province, Ecuador, one of a series obtained on 8 June 2011 by Elicio E. Tapia, Sofía Carvajal-Endara and Henry Grefa. GoogleMaps
Paratypes. ( Total 48: 10 adult males, 21 adult females, 17 juveniles). Ecuador: Azuay : CJ 1392–7 , collected with the holotype ; KU 120676 (female), 120683 –4, 120690, 120695 ( 4 juveniles), 120699 (male), 120705 (male), 120709 –10 (males), 120713 (female), 120718 –9 (female, male), 120721 (female), 120722 (male) from Cuenca , 2600 m, ( 02° 53' 57.91" S, 79° 01' 52.79" W), on 19 June 1968 by John D. Lynch GoogleMaps ; KU 138616 (female), 138619– 21 (male, females) from 4 km E: Cuenca 2540 m ( 02° 52' 59.88" S, 78° 22' 0.11" W), on 10 June 1970 by Thomas H. Fritts GoogleMaps ; KU 141572 (female) from 2.1 km S Cutchil , 2720 m ( 03° 06' 00"S, 78° 47' 59.99"W), on 16 May 1971 by Richard R. Montanucci GoogleMaps ; KU 141579 (female) from 3.5 km S Cutchil , 2785 m ( 03° 04' 59.87" S, 78° 47' 59.99" W), on 14 May 1971 by Richard R. Montanucci GoogleMaps ; KU 141582 (juvenile) from 8 km NW Cuenca , 2803 m ( 02° 51' 47.88" S, 78° 58' 59.88" W), on 16 May 1971 by Richard R. Montanucci GoogleMaps ; KU 141583 (female) from 8.8 km NW Cuenca , 3820 m ( 02° 52' 00.12" S, 79° 04' 00.12" W), on 15 May 1971 by Richard R. Montanucci GoogleMaps ; KU 203441 (male) from Laguna de Zurucuchu , 16 km NW Cuenca, 3200 m ( 02° 50' 23.99" S, 79° 07' 47.99" W), on 5 March 1984 by William E. Duellman GoogleMaps ; KU 129779–82 ( 4 females), 129783–91 ( 9 juveniles), 129793–94 (juveniles), 129795 (female), 129796 (juvenile) from Río Matadero , 12 km E Cuenca ( 02° 52' 59.88"S, 78° 58' 00.12" W), on 17 November 1968 by Craig E. Nelson. Cañar GoogleMaps : KU 141571 (male) and KU 141573 (female) from Biblián , 2620 ( 02° 42' 00" S, 78° 52' 00.12" W), on 17 and 23 May 1971, respectively, by Richard R. Montanucci GoogleMaps ; KU 142620–24 ( 4 females, male), from Biblián , 2620 m ( 02° 42' 00" S, 78° 52' 00.12" W), on 25 July 1971 by William E. Duellman GoogleMaps ; KU 147113 (female) from Biblián , 2620 m ( 02° 42' 00" S, 78° 52' 00.12" W), on 15 January 1972 by John E. Simmons GoogleMaps .
Referred specimens. (Total 36, 28 adult males, 8 adult females). Ecuador: Azuay: QCAZ 1239 View Materials (male), from Sigsig , 2480 m ( 03° 03' 08.4" S, 78° 48' 10.56" W), on 24 July 1989 by Luis A. Coloma and Luis E. López GoogleMaps ; QCAZ 26309 View Materials (female), from Cumbe , 2740 m ( 03° 05' 55.64" S, 79° 00' 28.73" W), on 8 August 2003 by Patricio Vargas GoogleMaps ; QCAZ 26348–50 View Materials (males), 26353 (female), 26357–64 ( 8 males), from Cumbe , 2740 m ( 03° 05' 55.64" S, 79° 00' 28.73" W), on 9 August 2003 by Luis A. Coloma, Ítalo G. Tapia, Andrés Merino Viteri, Erik Wild and Patricio Vargas Mena GoogleMaps ; QCAZ 26354 View Materials (female), from Cumbe , 2740 m ( 03° 05' 55.64" S, 79° 00' 28.73" W), on 9 August 2003 by Ítalo G. Tapia GoogleMaps ; QCAZ 26357–8 View Materials (males), from Cumbe , 2740 m ( 03° 05' 55.64" S, 79° 00' 28.73" W), on 9 August 2003 by Andrés Merino Viteri GoogleMaps ; QCAZ 31477 View Materials (male), from Sigsig , 2424 m ( 03° 03' 08.4" S, 78° 48' 10.56" W), on 8 March 2006 by Ítalo G. Tapia GoogleMaps ; QCAZ 31509 View Materials , 31511 View Materials (males), from Sigsig , 2424 m ( 03° 03' 8.4" S, 78° 48' 10.56" W), on 9 March 2006 by Ítalo G. Tapia and Giovanni Onore GoogleMaps ; QCAZ 34131 View Materials (female), from Cuenca , 2579 m ( 02° 53' 57.91" S, 79° 01' 52.79" W), on 27 February 1979 by Fernando I. Ortiz GoogleMaps ; QCAZ 37375–6 View Materials , 37379–81 View Materials , QCAZ 37386 View Materials ( 6 males), from unknown locality, on 31 May 2007 by Zoológico Amaru ; QCAZ 38232 View Materials (male), from Carmen del Guzho , 2666 m ( 02° 56' 00" S, 79° 03' 00" W), on 16 June 2007 by Hari González Maldonado GoogleMaps ; QCAZ 39384–5 View Materials (female, male), from Tarqui, Patapamba , 2600 m ( 03° 00' 38.16" S, 79° 01' 53.43" W), on 21 June 2007 by Ernesto Arbeláez Ortiz GoogleMaps ; QCAZ 42720–1 View Materials (males) from Cuenca , 2579 m ( 02° 53' 57.91" S, 79° 01' 52.79" W), on 3 October 2007 by Sofía Carvajal-Endara and Flavio Jaramillo GoogleMaps ; QCAZ 47106 View Materials (male), from Tarqui , 2741 m ( 03° 00' 57.17" S, 79° 02' 40.13" W), on 21 June 2007 by Ernesto Arbeláez Ortiz. Cañar GoogleMaps : QCAZ 42826 View Materials , 42835 View Materials , 42841 View Materials (females), from Papaloma de la Nube, 3011 m ( 02° 40' 21.54" S, 78° 54' 21.28" W), on 12–14 August 2008 by Sofía Carvajal-Endara GoogleMaps .
Diagnosis. Included in the genus Gastrotheca by having a closed brood pouch on dorsum of female. A moderately large species (49.0– 61.6 mm SVL in females, n = 28; 45.2–53.7 mm SVL in males, n = 38), with tibia length 39¯50% SVL, slightly shorter than length of foot; (2) interorbital distance slightly larger than width of upper eyelid; (3) skin on dorsum finely granular, not co-ossified with skull, lacking transverse ridges; (4) supraciliary processes absent; (5) heel lacking calcar or tubercle; (6) tympanic annulus weakly granular to smooth; (7) Fingers I and II about equal in length, width of discs greater than digits; (8) fingers unwebbed; (9) webbing between external toes extending to antepenultimate subarticular tubercle on Toe IV, to penultimate subarticular tubercle on Toe V; (10) in life, dorsum green, tan, brown, or reddish-brown with or without dark paravertebral marks; (11) head markings consisting of pale labial stripe and, in some individuals, an inconspicuous canthal stripe; (12) pale white or cream dorsolateral stripe present; (13) flanks tan, brown, or green with cream flecks or spots; groin and anterior and posterior surfaces of thighs translucent cream without marks and with a faint pale blue tinge in some; (14) venter uniform creamy white; (15) in females, brood pouch single, dorsal.
In comparison with similar species in Ecuador, G. cuencana is most like G. litonedis , G. lojana , G. elicioi , and G. riobambae . It differs from these species by having a uniform creamy white venter; whereas the venter is pale gray in G. litonedis , and cream with dark flecks, spots or mottling in G. lojana , G. elicioi , and G. riobambae . In G. cuencana the dorsal surfaces of fingers are cream, whereas in the other species dorsal surfaces of fingers are of the same color as the rest of the body (tan, brown, or green). Gastrotheca cuencana also differs from G. litonedis , G. elicioi , and G. riobambae by lacking a dark canthal stripe. Moreover, in G. cuencana the groin and anterior surfaces of thighs are mostly translucent cream without marks; whereas, in G. litonedis the groin and anterior surface of thighs are usually bronze-brown or tan with cream or darks flecks and spots. In G. lojana and G. riobambae the groin and anterior surfaces of the thighs are usually cream with black mottling; in G. elicioi the groin and anterior surfaces of thighs usually have dark and cream flecks and spots. In profile, the snout in G. cuencana is rounded above and inclined anteroventrally, whereas the snout in G. elicioi , G. lojana , and G. litonedis is bluntly rounded. Furthermore, G. cuencana has a conspicuous, elevated row of dorsolateral warts whereas these are barely raised in G. litonedi s; a narrow, cream supracloacal stripe is present in G. litonedis , G. lojana , and G. riobambae ; whereas the stripe is fragmented in G. elicioi and absent in most G. cuencana . Gastrotheca lojana and G. riobambae also differ from G. cuencana and G. litonedis by having a complex call, whereas the call in the latter two species is a simple call. Additionally, the call of G. cuencana has a lower note rate, shorter note duration, longer inter-note interval and a lower dominant and 90% bandwidth frequency compared with G. litonedis . Finally, G. cuencana differs from its sister species ( G. litonedis ) by having a genetic distance of 1.2% (in a DNA dataset of 438 bp, 16S gene).
Gastrotheca cuencana occurs in syntopy with G. pseustes ( sensu lato) throughout most of its range. Gastrotheca pseustes differs from G. cuencana by having coarsely granular skin on the dorsum, in contrast to the usually finely granular dorsum in G. cuencana . In G. cuencana the venter is uniform creamy white, whereas it is cream or white with dark flecks or spots in G. pseustes ; also the call of G. cuencana is simple, whereas in G. pseustes the call is complex.
Description of holotype. An adult male ( Fig. 2 View FIGURE 2 ); body moderately robust; SVL 50.6 mm; head wider that long; snout rounded in dorsal view, rounded and inclined anteroventrally in profile; canthus rostralis round in section; loreal region concave, lips rounded; top of head flat; interorbital distance 104% of width of upper eyelid; internarial area slightly elevated; nostrils not protuberant, directed anterolaterally, posterior to level of anterior margin of lower jaw; diameter of eye greater than its distance from nostril; tympanum round, separated from eye by distance about equal to diameter of tympanum; tympanic annulus and membrane weakly granular; supratympanic fold moderately weak, extending from behind the tympanum to the insertion of the forelimb. Dentigerous vomerine processes narrowly separated medially, each bearing four teeth.
Arm robust; ulnar tubercles absent; hand and fingers moderately large (TFL 35% of SVL), unwebbed, with distinct narrow lateral fringes; discs moderately large and rounded, width of disc of Finger III greater that diameter of tympanum; relative lengths of fingers I=II<IV<III; subarticular tubercles prominent, round, conical in lateral profile, non bifid; supernumerary tubercles, small, numerous, conical; palmar tubercle absent, prepollical tubercle elliptical. Hind limb robust; tibia length 46% SVL; foot length 49% SVL; calcar, tarsal tubercles, and inner tarsal fold absent; outer metatarsal tubercle absent; inner metatarsal tubercle large, elliptical; toes moderately long; relative length of toes I<II<III=V<IV; basal webbing between Toes I and II; webbing formula for other toes II1— 2III 1— 2IV 2— 1V; subarticular tubercles moderately large, rounded; supernumerary tubercles, numerous, minute and rounded. Nuptial pad absent.
Skin on dorsum finely granular; skin on flanks coarsely granular; skin on throat, venter surfaces of thighs, and arms granular; skin on belly areolate; skin on venter surface of shanks smooth; numerous small tubercles lateral to cloacal opening. Vocal sac single, median, subgular. Vocal slits present at posterior lingual margins of mandibles. Tongue broad, suboval, not notched posteriorly, fully attached to mouth floor.
Coloration in life ( Fig. 2 View FIGURE 2 ). The dorsum is yellowish brown with two darker, black delineated, continuous, paravertebral markings. These markings extend to the sacrum where they bifurcate and become two rows of irregular blotches at each side of the vertebral axis. The dorsal surfaces of the limbs are yellowish brown with irregular gray blotches. Cream dorsolateral and labial stripes are present; the latter continues to the insertion of forelimb; a faint brown canthal stripe is present. The flanks are bronze-brown with small black spots. The venter, groin, and anterior surfaces of the thighs are uniform creamy white; the posterior surfaces of the thighs have a faint bluish coloration. The dorsal surfaces of the fingers are cream. The gular region is creamy gray laterally. The iris is reddish bronze with black reticulations.
Coloration in preservative. The dorsum is tan with two well-defined, gray paravertebral markings, which bifurcate at the level of sacrum to become two rows of irregular blotches on each side of the vertebral axis. A white dorsolateral stripe is present. The dorsal surfaces of limbs are tan with irregular gray blotches. A cream labial stripe continues to the insertion of the forelimb; a faint canthal is present. The flanks are gray; the venter, groin, and anterior surfaces of thighs are uniform creamy white; the posterior surfaces of thighs have a faint bluish coloration. The gular region is creamy gray.
Measurements (in mm). SVL: 50.4, TIBL: 21.9, FL: 24.9, HL: 16.9, HW: 19.1, IOD: 5.2, EW: 5.0 IND: 3.3, ED: 5.6, EN: 4.1, TD: 2.8, FFL: 10.0, TFL: 17.5, TFD: 3.4.
Variation. Morphometric variation of 28 females and 38 males is summarized in Table 3. Females are larger than males (54.2± 2.9 mm; 49.6± 2.4 mm). The skin on the dorsum is finely granular in most, but it varies in some specimens from weakly areolate to coarsely granular. All adults have a supratympanic fold that usually extends from the posterior part of the tympanum to point above the insertion of forelimb. The tympanic annulus is usually smooth, but some individuals (e.g., CJ 1395, 1940) have a slightly granular annulus ( Fig. 3A, C View FIGURE 3 ). Each dentigerous vomerine process has 2–7 teeth (4.4±1.1, n = 39).
Color variation in preservative. Preserved specimens have bluish gray dorsum (tan and reddish-brown in CJ 1397). Faint paravertebral marks are present. A canthal stripe is absent in most specimens; the flanks and posterior surfaces of the thighs are dark gray with white spots evenly distributed in some specimens. In all specimens, a prominent white labial stripe extends from the posterior margin of the lip to the insertion of the forelimb. A narrow white dorsolateral stripe is evident in most specimens. The groin and the anterior surfaces of the thighs are uniform creamy gray. The ventral surfaces are uniform creamy white. The posterior surfaces of thighs usually are creamy gray, but they are pale blue in some specimens. Some males have yellowish brown nuptial pads on the medial surface of the thumb.
Color variation in life. ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ). The dorsum is uniform green (CJ 1393), reddish brown (CJ 1397), or tan (CJ 1392) with (CJ 1391) or without (CJ 1392) darker paravertebral marks. When present, the paravertebral markings usually are well defined; at the level of the sacrum they become two rows of irregular blotches at each side of the vertebral axis. The ventral surfaces are uniform creamy white. A cream labial stripe is extends from the posterior margin of the lip to the insertion of the forelimb; a dark brown canthal stripe is absent in most individuals. The tympanum is brown, tan, or olive green. The iris is reddish bronze with black reticulations. A creamy white dorsolateral stripe is evident in most individuals. The flanks are cream, bronze brown, or green. The groin and anterior surfaces of the thighs are translucent cream without dark markings ( Fig. 3 View FIGURE 3 ). The posterior surfaces of the thighs are translucent, cream, or pale blue or green. When present, supracloacal and heel stripes are cream or tan. The ventral surfaces of the shanks have a faint tinge of pale blue. The gular surface varies from white to creamy gray.
Tadpoles. Tadpoles belong to Type IV tadpole of Orton (1953), and the exotroph, benthic ecomorphological guild. All measurements are expressed in millimeters. The following description is based on a specimen in Stage 38 (CJ 1945a), from a series of eight tadpoles (CJ 1945) obtained from a pond at south Cuenca, 2579 m, Azuay Province, Ecuador, by Elicio E. Tapia, Sofía Carvajal-Endara, and Henry Grefa on 8 June 2011 ( Fig. 5A View FIGURE 5 ). Total length 44.6; body length 17.3 (39% of total length). Body ovoid in dorsal and lateral views, slightly depressed; throat slightly concave in lateral profile, sloping from tip of snout to belly; body width at the level of spiracle 11.5, and height at same position 9.3, head width at level of eyes 8.5. Lateral line system present but barely visible, supraorbital and infraorbital lines both rising at level of tip of snout, extending parallel to the eye and making contact immediately behind eye; angular line descending vertically just porterior of eye to throat, it dorsally contacts with post infraorbital line; anterior oral line descending vertically from oral disc level and behind nares level to throat; it makes a curve that parallels infraorbital line, forming a circuit continuous with angular and loreal lines; dorsal body and middle body lines not visible. Nostril medium sized (in proportion to body length), ovoid, protruding, having a fleshy annulus, its opening directed anterolaterally. Snout–nostril distance 2.3; internarial distance 2.4. Eye directed dorsally; eye length 1.8, eye width 1.8; interorbital distance 4.3. Spiracle sinistral, located at midbody level, spiracular opening oriented posteriorly; distance from tip of snout to spiracular opening 11.0; end of spiracular tube rounded, attached to body wall, inner wall of spiracular tube not evident; tube length 3.1, tube transverse width 2.0. Vent tube dextral, opening oriented posteriorly, tube length 1.7, tube transverse width 1.8. Tail length 27.2; caudal musculature slender, narrowing gradually until tail terminus; caudal muscle height 3.7, width 3,2; caudal fins well developed and proportional, arising near tail-body junction, forming a low hump; dorsal fin height 3.0, ventral fin height 2.8; maximum height of tail 9.3; tail terminus rounded, caudal musculature not reaching fin terminus.
Oral disc small, ventral, located near tip of snout, not protruding laterally beyond body; transverse width 4.7. It is surrounded by a uniserial row of marginal papillae, interrupted medially on upper lip; lower lip papillae alternate in orientation, giving appearance of two rows; upper lip with 29 papillae on right side and 27 papillae on left side; lower lip bearing 62 marginal papillae; upper jaw sheath medium-sized, forming a smooth arch and finely serrated, height 0.36, transverse width 2.8 (60% of width of oral width); lower jaw sheath V- shaped, open and finely serrated, width 1.92, height 0.7. Labial tooth row formula 2/3(1); tooth rows lengths: A1: 3.6, A2: 3.5, P1 right row 1.5, P1 left row 1.6, P1 gap 0.2, P2: 3.4, P3: 3.0. ( Fig. 6B View FIGURE 6 ).
Color in preservative. Dorsum dull gray with darker areas on flanks, above eye, and on throat; body contour and snout translucent. Caudal musculature and fins with scattered cream dots, with higher suffusion near tail base, otherwise translucent. Venter pale, translucent in belly region, guts exposed; eyes lavender gray; oral apparatus translucent.
Color in life. In dorsal view, body olive-cream; snout and flanks paler olive-cream. In ventral view, guts dark gray in belly area; reddish gills visible throughout throat. Caudal musculature covered by a nearly continuous line of cream marks, proximal half of musculature reddish cream with myomeres and medial line well defined, distal half translucent; dorsal and ventral fins translucent, with scattered cream dots, heavily suffused near tail-body junction. Spiracle, oral apparatus, and legs olive-cream. Iris gold.
Variation. Variation of 28 meristic characters of tadpoles in Stages 37–39 (CJ 1945) are shown in Table 4. Total length varies between 42.3 (Stage 37) and 48.7 (Stage 39) and tail length proportion remains about 61%; labial tooth row formula is 2/3(1). The number of marginal papillae varies among specimens and Gosner stages, variation in number of ventral papillae at lower lip is high (39–64).
We documented changes in coloration during ontogenetic development of one, mostly brown individual (CJ 1945) ( Fig. 7 View FIGURE 7 ). At Stage 41, the dorsum and flanks are diffuse brown with a well-defined pattern of brown-gray paravertebral stripes and a couple of blotches at posterior dorsolateral body. A dark, gray-black stripe borders the canthal and dorsolateral body extending to level of midbody, bordered dorsally by a creamy-white stripe that becomes wider at the distal end. The iris has a red suffusion. By Stage 46, markings on the dorsum and flanks are better defined and the black-gray stripe of flanks becomes a nearly black stripe; fingers are yellowish cream. Scattered green pigments are present in the anterior portion of the paravertebral stripes.
Comparisons. Tadpoles of Gastrotheca cuencana may occur in sympatry with the ones of G. pseustes 2. Gastrotheca cuencana differs from it by having less pigmentation on dorsum and venter, by lacking a reticulated pattern on flanks and by having a flesh colored tail musculature.
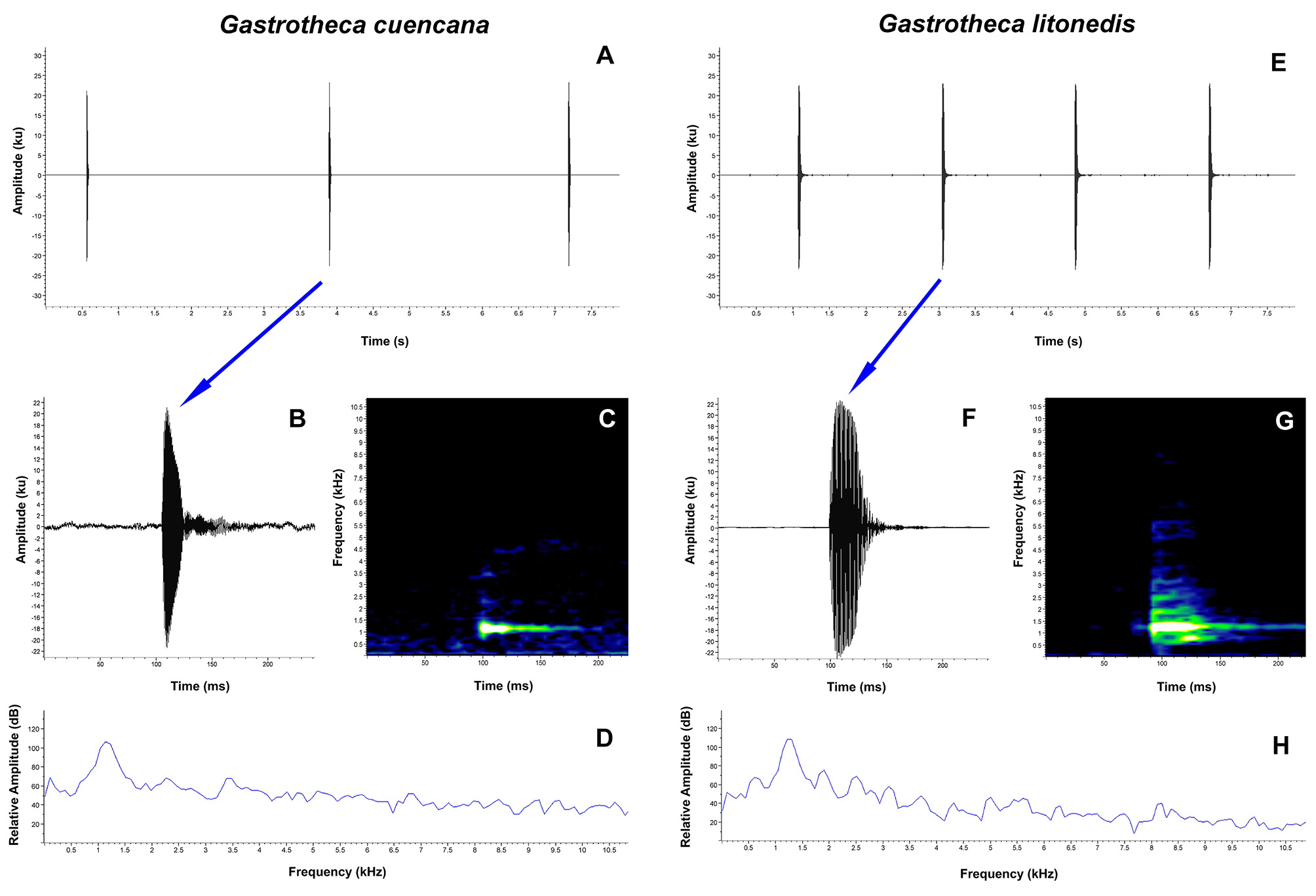
Vocalization. Eight individuals of Gastrotheca cuencana were recorded from two locations in Azuay Province (3 individuals from Cuenca and 3 from Río Mazán) and from one location in Cañar Province (2 individuals from Papaloma de la Nube) (Appendix III). Descriptive statistics of the acoustic variables are provided in Table 5. The advertisement call of G. cuencana is a simple call, composed of a series of short, single-pulsed notes, emitted at regular intervals ( Fig. 8 View FIGURE 8 A–D). In our recordings the calls have between 3 and 19 notes per call. The notes had a mean duration of 0.016 s (SD = 0.007) and a mean inter-note interval of 2.783 s (SD = 1.004). The mean dominant frequency of the call was 1147.9 Hz (SD = 99.536), with a mean 90% bandwidth of 1018.6–1269.4 Hz. The fundamental frequency and harmonics are not clearly recognizable.
……continued on the next page
Comparisons. The advertisement call of Gastrotheca cuencana is similar to that of G. litonedis , all the other species of Gastrotheca in the southern Ecuadorian Andes have complex calls. However, the call of G. cuencana has a lower note rate, shorter note duration, longer inter-note interval and a lower dominant and 90% bandwidth frequency compared with G. litonedis ( Table 5).
Distribution and ecology. Gastrotheca cuencana is known from the Paute Basin, which includes the eastern and western Cordilleras ( Fig. 9 View FIGURE 9 ). Its elevational range is 2407–3172 m in an area of extent of occurrence of about 1168 km 2.
This nocturnal, semiarboreal species inhabits mostly disturbed areas and a few forests in the Evergreen High Montane Forest of the Cordillera Occidental of the Andes (Ministerio de Ambiente del Ecuador 2012), where the average annual rainfall is 660–1122 mm and the average annual temperature is 9.6–15.2 °C ( Fick & Hijmans 2017). This species is found in pasture lands and urban grasslands where lentic water is available for tadpoles; thus, the frogs occur close to irrigation ditches, water reservoirs, temporary ponds or wetlands. The holotype and eight other individuals were collected in grassland in an urban area (Cuenca). Males were calling from small shrubs at approximately 80–100 cm above the ground. Females were perched on branches of shrubs at approximately 40 cm above the ground and also among the grass. A brooding female is depicted in Figure 10K View FIGURE 10 . Eight tadpoles were found in a small temporary pond. Gastrotheca cuencana is syntopic with G. pseustes ( sensu lato) throughout most of its range.
Conservation status. We suggest that Gastrotheca cuencana should be considered as Endangered according to criteria B1ab(i,ii,iii,iv) of the IUCN Red List. Its small area of known occurrence ( 1168 km 2) is fragmented, and its habitats are in heavily human populated areas. For example, the city of Cuenca is rapidly growing and occupying previous rural areas ( Fig. 11B View FIGURE 11 ) where swampy areas are being drained. One population inhabits Cajas National Park, which affords protection from habitat destruction. Some efforts ( in situ and ex situ) to protect this species are underway by parties of the Cajas National Park, by Amaru Zoo ( Amphibian Ark 2013), and the city council of Cuenca ( Siavichay-Pesántez et al. 2016). However, these efforts urgently need to be reinforced.
Etymology. The specific name cuencana is an adjective referring to people born in Cuenca, the capital city of the province of Azuay, where this species occurs. We also use this name to highlight the pioneer efforts taking place in Cuenca, throughout the project Conservation of the Urban Biodiversity of the Municipality. The project is directed to protect the urban and suburban species of frogs through biorestauration and protection of watersheds, river banks, wetlands, and frog ponds ( Arbeláez Ortiz & Vega Toral 2008, Siavichay-Pesántez et al. 2016).
Comments. Gastrotheca cuencana was confused with G. litonedis in the past. Our recognition of both species emphasizes the discrete morphological and acoustic differences between them. Gastrotheca cuencana belongs to the subgenus Duellmania ( sensu Duellman 2015) .








| KU |
Biodiversity Institute, University of Kansas |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |