
Arktocara yakataga, Alexandra T. Boersma & Nicholas D. Pyenson, 2016
|
publication ID |
https://doi.org/ 10.7717/peerj.2321 |
|
publication LSID |
lsid:zoobank.org:pub:0194A593-DBE0-47CA-A41F-04A37931BA2F |
|
DOI |
https://doi.org/10.5281/zenodo.5658608 |
|
persistent identifier |
https://treatment.plazi.org/id/FBCF0EAA-7BBB-4EF0-8186-7548993098D1 |
|
taxon LSID |
lsid:zoobank.org:act:FBCF0EAA-7BBB-4EF0-8186-7548993098D1 |
|
treatment provided by |
Plazi |
|
scientific name |
Arktocara yakataga |
| status |
sp. nov. |
Arktocara yakataga , sp. nov.
( Figs. 2 10 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 and Table 1 View Table 1 )
LSID: urn:lsid:zoobank.org:act:FBCF0EAA7BBB4EF081867548993098D1
Holotype. USNM 214830, consisting of an incomplete skull lacking the rostrum anterior of the antorbital notches, tympanoperiotics, dentition and mandibles (see Fig. 2 View Figure 2 ).
Type locality. The precise geographic coordinates for the type locality of Arktocara yakataga are unknown. The type specimen ( USNM 214830) was discovered and collected in 1951 by United States Geological Survey ( USGS) geologist Donald J. Miller (1919 1961), who was mapping what was then the Yakataga District of Alaska (now the Yakutat City and Borough) between 1944 and 1963. Archival notes housed with the specimen at USNM state that Miller found the specimen in the Poul Creek Formation within the thenYakataga District (see Age, below). Therefore, we delimit the area for the type’s provenance to exposures of the Poul Creek Formation in the Yakutat City and Borough, Alaska, USA, in a grid ranging approximately from 60 22 0N, 142 30 0W to 60 0 0 0N, 143 22 0W (see Fig. 1 View Figure 1 ). While the formation has been named from its exposures along Poul Creek, it has been suggested that the most abundant macrofossils from this unit have been collected from outcrops along Hamilton Creek, White River, and Big River near Reare Glacier ( Taliaferro, 1932). It is possible that Miller collected USNM 214830 from one of these exposures. Formation. Poul Creek Formation.
Age. Archival documentation accessioned in the Department of Paleobiology with USNM 214830 indicate that the type specimen was collected from an unknown locality exposed about 400 500 m below the top of the Poul Creek Formation, which has a total stratigraphic thickness of around 1.9 km ( Plafker, 1987). The Yakutat terrane of Southeast Alaska consists of the Kulthieth, Poul Creek, and Yakataga Formations ( Perry, Garver & Ridgway, 2009; Plafker, Moore & Winkler, 1994; Miller, 1971). The Kulthieth Formation consists of mostly organicrich sandstones deposited in nonmarine alluvial, deltaic, barrier beach and shallow marine environments, and is Early Eocene to Early Oligocene (54 33 Ma) in age based on the fossil assemblages present ( Perry, Garver & Ridgway, 2009). The Upper Eocene to possibly Lower Miocene (40 20 Ma) Poul Creek Formation conformably overlies the Kulthieth Formation ( Plafker, 1987; Miller, 1971). It is estimated to be approximately 1.9 km thick, and is composed of siltstones and organicrich sandstones, in part glauconitic recording a marine transgression, interrupted by deposits of the Cenotaph Volcanics ( Plafker, 1987). Finally, unconformably overlying the Poul Creek Formation is the Miocene to Pliocene Yakataga Formation ( Miller, 1971). It is composed mainly of tillite and marine strata ( Perry, Garver & Ridgway, 2009).
The Poul Creek Formation itself is broadly constrained to approximately 40 20 million years in age, from the latest Eocene to possibly early Miocene in age ( Plafker, 1987; Miller, 1971). The depositional age of the unit has been further constrained to 24 to 29 Ma, or a mid to late Oligocene age, based on detrital zircon fissiontrack analyses of young grainage populations ( Perry, Garver & Ridgway, 2009). Using the broadest time duration for the formation (20 million years) and the coarse stratigraphic thickness of the sediments within it (2 km), a constant rate of sedimentation would suggest that the stratigraphic position of USNM 214830 at 500 m below the top of the formation would be roughly equivalent to an geochronologic age of 25 million years, an estimate that is consistent to the detrital zircon analyses. Overall, we propose a late Oligocene, or Chattian age for Arktocara, although we cannot exclude a Rupelian antiquity.
Diagnosis. Arktocara is a small to medium sized platanistoid odontocete (approximately 2.26 m in total length), which belongs, based on one equivocal synapomorphy, to the nodebased Platanistoidea: width: width of the premaxillae>50% of the width of the rostrum at the antorbital notch (character 51[1]). More convincingly, Arktocara belongs to Platanistoidea based on its affinities to other members of the Allodelphinidae that possess unequivocal synapomorphies of the Platanistoidea (see ‘Discussion’ for further comments on the relationship of Allodelphinidae within the Platanistoidea). We also note that, for the purposes of this diagnosis, we use a broad definition of Waipatiidae that includes Otekaikea spp. (see Tanaka & Fordyce (2015a)), and Squalodelphinidae sensu Lambert, Bianucci & Urbina (2014) . See ‘Discussion’ for further comments on systematics of these groups.
Arktocara can be differentiated from all other platanistoids by the following combination of character states. Arktocara differs from Waipatiidae and Squalodelphinidae in having: a postglenoid process of squamosal greatly reduced (character 114[1]); an occipital shield bearing a distinct sagittal crest (character 118[1]); length of the zygomatic process as percent of the greatest width of the maxillae across the postorbital processes <30% (character 152[1]); lacking a dorsal condyloid fossa (character 119[0]); and lacking any asymmetry in the vertex (character 98[0]).
Arktocara also differs from Waipatiidae and Platanistidae in having: frontals posterior to the nasals and between the premaxillae wider than the maximum transverse width across the nasals (character 95[0]); and lacking an anterior transverse ridge and large tympanosquamosal recess, with middle sinus still inferred to be present (character 144[1]). Arktocara further differs from Waipatiidae in having: a lacrimal that wraps around the anterior edge of the supraorbital process of frontal and slightly overlies its anterior end (character 37[0]); maxilla forming the dorsolateral edge of the internal opening of the infraorbital foramen (character 43[0]); ventromedial edge of the internal opening of infraorbital foramen formed by maxilla and palatine (character 44[1]); a nuchal crest weakly convex anteriorly in dorsoposterior view (character 117[1]); a lateral end of the groove for the mandibular branch of the trigeminal nerve wrapping laterally around posterior end of pterygoid sinus fossa and opening primarily anteriorly (character 148[0]); the angle formed by the basioccipital crests in ventral view between 15 40 (character 157[1]); and in lacking a premaxillary crest or posterior maxillary crest adjacent to the nasal (character 72[0]).
Arktocara differs from all Platanistidae and Squalodelphinidae in having the anterolateral corner of the maxilla overlying the supraorbital process of frontal being thin and of even thickness to parts posteromedial (character 64[0]). Arktocara also differs from
all Platanistidae in having: the apex of the postorbital process of frontal projected
posterolaterally and slightly ventrally (character 46[0]); the ratio of the greatest width of the premaxillae to the greatest width of maxillae at the level of postorbital processes between 0.49 0.38 (character 76[1]); a shallow emargination of the posterior edge of zygomatic process by the sternomastoid muscle fossa in lateral view (character 111[1]); the width of the squamosal lateral to the exoccipital in posterior view as a percent of the greatest width of the exoccipitals <15% (character 112[0]); fossae for the preorbital lobe of the pterygoid sinus present in the orbit (character 134[1]); shallow posterior portion of the periotic fossa of the squamosal (character 151[1]); posteroventralmost point on the basioccipital crest forming a closely appressed flange with a narrow crease separating exoccipital dorsally from the rest of crest (character 156[1]); and lacking a pneumatic maxillary crest overhanging medially (character 65[0]). Finally, Arktocara differs from all Squalodelphinidae in having: a narrower width of the premaxillae at the antorbital notches as a percent width of the rostrum at the antorbital notch (50 64%) (character 51[1]).
Arktocara differs from all other Allodelphinidae in having: a reduced postglenoid process of the squamosal (character 114[1]); and the posteroventralmost point of the basioccipital crest forming a closely appressed flange separating the exoccipital dorsally from the rest of the crest by a narrow crease (character 156[1]). Arktocara differs from Allodelphis pratti and Goedertius oregonensis in having: both premaxillae extending posterior to the nasals (character 58[1]); and the ratio of the greatest width of the premaxillae to greatest width of the maxillae at the level of the postorbital processes between 0.49 0.38 (character 76[1]).
Arktocara also differs from Goedertius oregonensis and Zarhinocetus errabundus in having: the width of the premaxillae between 50 64% of the width of the maxillae a the level of the antorbital notches (character 51[1]); a uniformly thin anterolateral corner of the maxilla overlying the supraorbital process of the frontal (character 64[0]); length of the zygomatic process of the squamosal 30% of the width of the maxillae at the postorbital processes (character 152[1]); lacking a rostral basin (character 50[0]), lacking a posterior dorsal infraorbital foramen placed posteromedially near the posterior extremity of the premaxilla (character 60[0]); lacking a dorsal condyloid fossa (character 119[0]) and having a symmetrical cranial vertex (character 98[0]).
Arktocara also differs from Allodelphis pratti and Zarhinocetus errabundus in having: two anterior dorsal infraorbital foramina (character 49[1]); and a shallow emargination of the posterior edge of the zygomatic process by the sternomastoid muscle fossa in lateral view (character 111[1]). Arktocara further differs from Goedertius oregonensis having: a fused lacrimal and jugal (character 39[1]); the apex of the postorbital process of the frontal directed posterolaterally and slightly ventrally (character 46[0]); a triangular or anteroposteriorly widened falciform shaped postorbital process of the frontal (character 47[2]); one posterior dorsal infraorbital foramina of the maxilla (character 59[1]); nuchal crest weakly convex anteriorly in dorsoposterior view (character 117[1]); and the posterior edge of the vomer terminating on the basisphenoid (character 154[0]).
Arktocara further differs from Zarhinocetus errabundus in having: the transverse width of the nasal>70% the length of the nasal (character 91[2]); a distinct sagittal crest on the occipital shield (character 118[1]); and in lacking an anterior transverse ridge and large tympanosquamosal recess (character 144[1]).
Lastly, Arktocara displays the following apomorphies: a weakly developed antorbital notch (character 10[0]); straight lateral margin of the right premaxilla posterior to premaxillary foramen (character 56[1]); a Ushaped nasal frontal suture (character 94[2]); and a greatly reduced postglenoid process of the squamosal (character 114[1]).
Etymology. The species epithet ‘yakataga’ derives from the Tlingit name for the point of land along the southeast coast of Alaska between modern day Kayak Island and Ice Bay. This point, currently called Cape Yakataga, is located directly southwest of Watson Peak and represents the southeastern boundary of a floodplain drained by the Bering Glacier. The name Yakataga was first published by Tebenkov (1852: map 7), who was a cartographer and hydrographer of the Imperial Russian Navy, as ‘‘M[ys] Yaktaga’’ on an 1849 map of Alaska. The geographic place name has been alternatively spelled Cape Iaktag, Cape Yakaio, Cape Yakatag, and Yokataga Reef ( Orth, 1967). According to the Geographic Names Information System (GNIS, 2016), developed by USGS in cooperation with the United States Board of Geographic Names (BGN), the name ‘‘Yakataga’’ means ‘‘canoe road,’’ referring to two reefs that form a canoe passage to the shore of the village.
Description
Anatomical terminology follows Mead & Fordyce (2009). In most cases, description of individual elements derives from the most informative side of the skull, in terms of preservation; we note any morphological asymmetry if present. Besides overall erosion of the bone surfaces and loss of some skull elements, there appears to be no significant burialrelated distortion of the skull.
Skull
The holotype of Arktocara yakataga (USNM 214830) consists of an incomplete skull, measuring 23 cm in preserved length. The majority of the rostrum is missing, with an asymmetric transverse break approximately 2 5 cm anterior to the antorbital notch. The skull also lacks both nasals, jugals, tympanoperiotics, and the right occipital condyle (see Figs. 2 View Figure 2 and 3 View Figure 3 ). Most of the paroccipital processes of the exoccipitals are gone, large portions of the palatines and pterygoids are missing, small fragments along the lateral margins of the frontals and maxillae are incomplete, and the general condition of many osteological elements in the skull are poorly preserved. The skull may have been both mechanically and chemically prepared in the past (with no known documentation), including acid preparation, which may have contributed to the poor state of preservation for the osteological surfaces of many elements. Portions of the skull are obscured by a finegrained grey matrix of siltstone, especially infilling the mesorostral canal, the bony nares, the recesses of tympanoperiotic region, and the braincase (which is exposed via the foramen magnum and fenestrae in the supraoccipital).
In dorsal view, the preserved skull is roughly hexagonal in overall shape ( Fig. 2 View Figure 2 ). The external nares are vertically oriented, and positioned at a level between the antorbital and postorbital processes. The vertex is particularly tablelike and square, composed of frontals, premaxillae, and nasals (missing). The parietals appear to be narrowly exposed on the vertex immediately anterior to the nuchal crest, between the frontals and supraoccipital whether this is natural, or an ontogenetic characteristic, is unclear. The vertex is bordered anteriorly by the externals nares, laterally by the maxillae and posteriorly by the nuchal crest of the supraoccipital. The nuchal crest is straight along the posterior edge of the vertex, but begins to curve posterolaterally as it approaches the temporal crest. The temporal fossae are visible in dorsal view due to an intertemporal constriction just anterior to the level of the nuchal crest, and the fossa is floored by a narrow valley (the squamosal fossa) between the squamosal plate and supramastoid crest.
In lateral view, the profile of the skull gradually slopes upwards from the level of the antorbital notch to posterior of the nares, where it levels out on the vertex ( Fig. 4 View Figure 4 ). The nuchal crest is well defined, and about the same height as the frontals on the vertex. The orbit is shallowly rounded dorsally (4.9 cm in length), with the maxilla completely overlying the frontal on the thin supraorbital process, except on the postorbital process, where the frontal is exposed laterally. It is unclear whether the antorbital process of the frontal is completely covered by maxilla or not, but most of the medial antorbital process is composed of the lacrimojugal. The temporal fossa is trapezoidal in shape, with the temporal crest forming a right angle with the dorsal margin of the zygomatic process of the squamosal. The dorsal margin of the temporal fossa is roofed over by the frontal.
The preserved posterior portion of rostrum anterior of the antorbital notch is wide (8.8 cm anterior to the antorbital notch) and deep (6.5 cm at the level of the antorbital notch), with a broadly open and deep mesorostral canal (2.4 cm wide and 4.6 cm deep at the level of the antorbital notch). While there is material missing around the antorbital notch, the posterior margin of the right antorbital notch appears to be real, demonstrating it to be weakly developed, forming an Lshape with the lateral margin of the rostrum ( Fig. 2 View Figure 2 ). Though some of the anteromedial antorbital process is missing, not enough is missing to have drastically changed the shape of the antorbital notch region. In anterior view, the maxilla abruptly slopes upwards medially to the distinct premaxillamaxilla suture, and underlies the premaxilla along the entire preserved length of rostrum and likely the cranium ( Fig. 5 View Figure 5 ). The premaxilla therefore forms an anteroposteriorly elongated rectangular plateau surrounding the external bony nares, elevated in relation to the plane of the maxillae, appearing similar in transverse crosssection to a horst and graben system.
In ventral view, there is clearly a fossa for the preorbital lobe of the pterygoid sinus (a shallow depression surrounding the ventral infraorbital and sphenopalatine foramina), as well as for the hamular lobe of the pterygoid sinus (fossa anterior to the choanae). Also present are an extremely shallow tympanosquamosal recess for the middle sinus, and a middle pterygoid sinus fossa anterior to the periotic fossa ( Fig. 6 View Figure 6 ). There is not enough rostral material or paroccipital process to determine the presence of an anterior or posterior sinus respectively, and the presence of a peribullary sinus is difficult to ascertain.
Premaxilla
Both of the premaxillae are symmetrical, and overlie either the maxillae or the frontals for their entire preserved length ( Fig. 2 View Figure 2 ). In lateral view, the premaxilla thins slightly as it passes around the external nares, otherwise maintaining a relatively even thickness on the cranium ( Fig. 4 View Figure 4 ). The premaxilla also appears to thin anterior of the antorbital notch (especially in right lateral view), lowering to the same level as the maxilla instead of rising dorsally above it. However, in anterior view, it is evident that the ventral margin of the left premaxilla sinks ventrally into a medial trough created by the maxilla while the dorsal margin remains level, accounting for the apparent reduction in thickness ( Fig. 5 View Figure 5 ). Both premaxillae are broken anteriorly, the right further anterior than the left and missing some of its anterolateral edge. The premaxillamaxilla suture is clear in dorsal view along the entire lateral length of the premaxilla, as well as in anterior view at the transverse crosssection of the rostrum. In dorsal view, the lateral margin of the premaxilla is mostly rectilinear, widening only 0.5 cm from the rostral break to a level anterior to the nares. As it passes laterally around the nares, the premaxilla gently bows out laterally, with the medial edge retreating more than the lateral edge so that the total width is reduced (0.8 cm on the right premaxilla). Posterior to the nares, the lateral edge remains straight posteriorly, but the medial edge expands slightly medially, once again widening the premaxilla. On the left side of the skull, lateral and posterior of the external nares, a narrow ledge of the medial margin of the maxilla laterally borders the premaxilla, where the premaxilla is separated from the maxilla (possibly diagenetically).
On the vertex, the posterior termination of the premaxilla lies on the frontal as an asymmetrical, spatulate lobe, tapering posterolaterally and bordered medially by frontal and the nasal fossa of the frontal, and laterally the maxilla ( Fig. 2 View Figure 2 ). There is no division of the premaxilla into a posteromedial splint and posterolateral plate (a widespread characteristic amongst odontocetes). The premaxillary sac fossa, roughly at the level of the antorbital notch, is shallow with a rough surface. No premaxillary foramina or associated premaxillary sulci are visible on the preserved length of the skull.
Maxilla
Only a small portion of rostral maxilla remains, including part of the maxillary flange on the left side, and just over 5 cm of the body of this element on the right side. In anterior view, the maxilla preserved on the rostrum slopes dorsally from the lateral edge to the premaxillary suture ( Fig. 5 View Figure 5 ). The premaxillary suture is distinct and unfused. The preserved maxillary flange on the left side is thin and flat. In lateral view, the maxilla gradually increases in depth posteriorly until anterior of the nares, where it reduces in dorsoventral depth to a thin plate forming the facial portion and ascending process of the maxilla ( Fig. 4 View Figure 4 ). In dorsal view, the maxilla flares out laterally at the level of the antorbital notch, which is shallow and Lshaped. Posterior to the antorbital process is broad and relatively flat. The right side bears two infraorbital foramina: one immediately posteromedial to the antorbital notch, and one at the level of the nares. The left maxilla has three infraorbital foramina, all in a sagittal plane from immediately posteromedial of the antorbital notch to a level anterior of the nares. The posteriormost dorsal infraorbital foramen on either side is the largest foramen, with two sulci each one directed posterolaterally and the other posteromedially giving the foramen a vshaped appearance in dorsal view. In the facial region, the maxilla gradually curves dorsomedially from the supraorbital process to the premaxillary suture, and the facial fossa is essentially flat. The maxilla does not extend to the postorbital process, though the suture with the frontal on the postorbital process is unclear. The ascending process suddenly curves dorsomedially towards the lateral edge of the vertex, and the posteromedial margin of the maxilla curves dorsolaterally to terminate in a sharp triplepoint junction with the nuchal, temporal, and orbitotemporal crests.
In ventral view, the hard palate of maxilla curves dorsolaterally from the midline to the lateral edge, where it flattens out on the maxillary flange ( Fig. 3 View Figure 3 ). No alveoli are present in the preserved palatal surface of the maxillae. A small gap between the maxillae along the midline of the hard palate reveals a thin ridge of the vomer, approximately 2.6 cm long and a maximum of 1 mm wide. Just anterior to the level of the antorbital notch, a narrow palatine groove of the maxilla for vessels and/or nerves begins approximately 1.5 cm lateral of the midline, and curves posterolaterally to terminate ventral of the sphenopalatine foramen. Midway along the palatine groove is the posterior palatine foramen, and the posterior portion of the groove is partially roofed over by fragments of the palatine. The distal maxillopalatine suture would have been at some point anterior and lateral of the palatine groove, though the palatine is incomplete and is missing this portion. Between the palatine groove and the medial lamina of the pterygoid, the maxilla is overlain by fragments of the palatine, though the sutures between the two are extremely unclear ( Figs. 2 View Figure 2 , 4 View Figure 4 and 6 View Figure 6 ). Lateral of the palatine groove, the maxilla overlies the medial process of the lacrimal, and encircles both the ventral infraorbital foramen and the dorsal margin of the sphenopalatine foramen. Posterior of the foramina, the maxilla terminates in an abutment with frontal.
Frontal
In dorsal view, the frontals are mostly covered by the maxillae, with the exception of exposures on the postorbital processes and the vertex ( Fig. 2 View Figure 2 ). The postorbital processes are asymmetrical in lateral view, with a larger and more robust right postorbital process pointing ventroposteriorly and reaching within 0.7 cm of the tip of the zygomatic process ( Fig. 4 View Figure 4 ). The left postorbital process is shorter and more smoothly curved ventroposteriorly. Though more of the length appears to be preserved in the right postorbital process than the left, it is difficult to determine whether the asymmetry is real or preservational. Moreover, the dorsal rim of the right orbit is missing and heavily eroded into the supraorbital process, yielding an incomplete view of the orbit on this side of the skull. The frontalmaxillary suture is indistinct as it passes in an anteroposterior direction across the postorbital processes.
The frontalmaxillary suture is also indistinct along the lateral edge of the vertex, where the maxilla rises dorsomedially to the edge of the vertex’s tabular surface ( Fig. 2 View Figure 2 ). The sutures are posteromedially convex in dorsal view, on either side of the vertex, curving from the temporal crest to the posterior premaxillamaxilla suture. On the vertex, the frontals are exposed as a wide, flat surface. They are separated from supraoccipital posteriorly by a narrow exposure of the parietals which may an artifact of ontogeny. As such, it is possible that the frontals did contact the supraoccipital in older individuals of this taxon. The frontal exposure is bordered by the maxillae laterally, and the premaxillae and mesethmoid anteriorly. Between the premaxillae, the frontals bear a shallow fossa for the missing nasal bones (see ‘‘Nasal’’ for further description).
Lacrimojugal
Most of the medial margin of the antorbital process is composed of the lacrimal. In dorsal view, a narrow margin of the lacrimal emerges from beneath the maxilla as a thin plate along the lateral and anterior edges of the antorbital process ( Fig. 2 View Figure 2 ). In ventral view, the medial process of the lacrimal extends posteromedially towards the ventral infraorbital foramen, but is overlapped by the maxilla ( Fig. 3 View Figure 3 ). The styliform portion of jugal is mostly missing, save a small piece of the jugal base where it is fused to the lacrimal. The jugal base is transversely wide and anteroposteriorly narrow. The lacrimal is covered posteriorly by the frontal.
Nasal
Though both nasal bones are missing, the frontal bones bear a distinct fossa between the premaxillae on the vertex that indicates where the bones would have been ( Fig. 2 View Figure 2 ). The fossa is bordered anteriorly by the mesethmoid and laterally by the premaxillae. Based on the extent of the nasal fossa, the nasals were wider anteriorly than posteriorly (from 2.1 to 1.6 cm), and minimally 1.9 cm in length, but could have been longer if they overhung the bony nares, as seen in the holotype of Allodelphis pratti ( Fig. 7 View Figure 7 ) The height of the nasals cannot be estimated, considering that they rise above the frontal to varying degrees in similar taxa such as Allodelphis pratti. All codings for the nasals of Arktocara yakataga in the phylogenetic analysis were made from measurements of this nasal fossa of the frontals, considered the minimum extent of the nasals.
Vomer
At the broken rostral tip in anterior view, the mesorostral canal is deeply vshaped ( Fig. 5 View Figure 5 ). The crosssection reveals the damaged vomer to be extremely thin (<1 mm), and lining all sides of the mesorostral canal. Anterior to the nares, the maxilla is exposed dorsal of the vomer, so that it forms the dorsal edge of the mesorostral canal and the anterior wall of the external bony nares, similar to Tursiops truncatus ( Montagu, 1821) (based on USNM 504560). Anterior to the nares, the vomer also curves medially to form the medial wall of the external bony nares, and the lateral walls of the nasal septum. Posterior of the nares, the vomer is obscured in dorsal view by unprepared matrix.
In ventral view, the vomer is visible as a long, thin crest running down the midline of the hard palate ( Fig. 3 View Figure 3 ). Anterior to the antorbital notch, the vomer is mostly obscured by the maxillae, but emerges at the level of the antorbital notch as a vertical wall separating the choanae, slightly wider at its base and thinning to a sharp crest ventrally. Posterior of the nares, the crest flattens and the vomer flares out laterally, adhering to the basisphenoid. Its suture with the basisphenoid is indistinct, roughly straight transversely, between the posterior lamina of the pterygoid and immediately anterior to the basisphenoidbasioccipital suture.
Mesethmoid
The mesethmoid composes the bulk of the nasal septum, narrowing dorsally and flanked on either side by the thin lamina of the vomer. Behind each choana is a rounded bony protuberance, likely composed of the lateral wings of the mesethmoid (referred to as the ectethmoid by some authors, e.g., Mead & Fordyce, 2009) ( Fig. 2 View Figure 2 ).
Parietal
The parietals are visible in dorsal and lateral view in the temporal fossa, where they are in contact with the frontals under the temporal crest, the supraoccipital along the occipital border, and partially underlie the squamosal plate with a semicircular suture across the temporal fossa. All of the sutures are indistinct. The parietal forms the lateral wall of the braincase as a thin, laterally convex plate. As with the supraoccipital, both parietals in the temporal fossa are broken, with a small rounded window revealing the matrixfilled braincase ( Fig. 4 View Figure 4 ). As previously mentioned, the parietals in dorsal view are exposed as a anteroposteriorly narrow strip along the posterior margin of the vertex. In ventral view, the parietal is again visible in the periotic fossa; having passed under the squamosal to form the lateral wall of the braincase, it emerges medial to the squamosal in ventral view as small, slightly concave surface, just posterior to the foramen ovale ( Figs. 3 View Figure 3 and 6 View Figure 6 ).
Supraoccipital
The supraoccipital is broadly visible in dorsal view, contributing to the entire length of the nuchal crest. The supraoccipital and frontals are very slightly separated along the vertex by an exposure of the parietals sutured to the supraoccipitals. The supraoccipital is also in contact with the parietals along the occipital border ( Fig. 2 View Figure 2 ). Lateral of the vertex along the nuchal crest, the supraoccipital is in direct contact with the maxilla. The nuchal crest is straight medially, but begins to curve posteriorly as it approaches the temporal crest. Along the parietal margin, the supraoccipital is a thin plate, with the edge oriented posterolaterally as it curves around the posterior edge of the temporal fossa. In posterior view, the supraoccipital is rectangular in shape ( Fig. 5 View Figure 5 ). A prominent external occipital crest divides supraoccipital sagittally, from the midpoint of the nuchal crest to the opisthion. On either side of the crest, the supraoccipital is very thin and slightly concave. Both these surfaces are broken into fenestrae, with rounded margins that reveal openings filled with matrix in the braincase. There is no evidence of a dorsal condyloid fossa of any significance. The contact of the supraoccipital with the exoccipital is indistinct, except around the foramen magnum, where the nuchal tubercle of the supraoccipital clearly tucks underneath the dorsal portion of the occipital condyle.
Exoccipital
Both exoccipitals are incomplete, missing all or part of the occipital condyle, and most of their ventral portions ( Fig. 5 View Figure 5 ). The supraoccipital suture is indistinct, but the contact with the squamosal is clear, along the posteroventral temporal crest, and on the ventral side of the skull. The exoccipital is thin along the lateral margin posterior of the temporal crest, thickening ventrally. The broken remains of the occipital condyles are sufficient to observe their robust size and width, composing approximately 70% of the total combined width of the exoccipitals. Only the dorsal portion of the left condyle remains. Its surface is smooth, posteriorly curved and laterally broad. The foramen magnum is elliptical in shape, almost twice as wide as it is tall (2.9 cm wide, 5.8 cm tall). Both ventral and dorsal condyloid fossa are very shallow and illdefined. Though both exoccipitals are missing most of their ventral portions, including the jugular notches and paroccipital processes, the left exoccipital does bear a small foramen that may represent the hypoglossal foramen, immediately lateral to the posterior end of the basioccipital crest. The exoccipital also forms a small knob separated from the posterior end of the basioccipital crest by a narrow cleft, possibly a vestigial medial crest of the exoccipital.
Basioccipital
In ventral view, the basioccipital widens posteriorly from 6.2 cm wide at its suture with the basisphenoid, to 8 cm at the posterior end of the basioccipital crest ( Fig. 3 View Figure 3 ). The element is ventrally concave, with basioccipital crests oriented laterally from a sagittal plane, and at an angle of approximately 12 degrees from the midline (opening posteriorly). The tympanic plates are thin where they overlie the basisphenoid, increasing in width posteriorly before tapering slightly and rounding off at their posterior ends. The right side of the posterior basioccipital crest is missing, though the left side is complete. The posterior end of the basioccipital crest is interrupted by a narrow cleft that separates a small knob from the rest of the tympanic plate. This small knob is immediately medial to the hypoglossal foramen (on the left side), and presumably the jugular notch. Though consistent with the rest of the basioccipital crest, the knob is actually formed by the exoccipital a condition described in Fordyce’ s (1994) description of Waipatia maerewhenua, and also seen in a number of other platanistoids, including Allodelphis pratti ( Figs. 7 9 View Figure 7 View Figure 8 View Figure 9 ). The suture with the basisphenoid, along the anterior margin of the basioccipital, is represented by a wavy margin near the midline. The suture is increasingly less distinct laterally, where the basioccipital extends anteriorly, overlapping the lateral margins of the basisphenoid and bordering the posterior lamina of the pterygoid along its medial edge. There is no strong evidence of a muscular tubercle for the insertion of the ventral rectus capitis muscle.
Sphenoid
The basisphenoid is visible on the ventral side of the skull, though it is mostly obscured by the basioccipital and vomer ( Fig. 3 View Figure 3 ). The basioccipital crests extend anteriorly to cover the lateral portions of the basisphenoid, completely obscuring any view of the contact between the basisphenoid and alisphenoid. The posteroventral plate of the vomer obscures the anterior margin of the basisphenoid, and spreads over the basisphenoid’s medial section to terminate immediately anterior to the wavy basioccipital suture, which appears partially fused. In ventrolateral view, the sphenoid reemerges from beneath the basioccipital, with the ventral carotid foramen tucked under the dorsolateral margin of the basioccipital crest ( Fig. 6 View Figure 6 ). A small portion of the basisphenoid is visible, wrapped laterally around the ventral carotid foramen. Anterolateral of the foramen, the alisphenoid extends laterally anterior to the periotic fossa as a thin plate. The alisphenoid passes anterior to the foramen ovale, and bears a long, thin groove for the mandibular nerve, extending anteriorly from the foramen ovale to the anterior margin of the alisphenoid, medial of the subtemporal crest of the squamosal. The alisphenoid continues to extend lateral of this groove, posteriorly contacting the medial edge of the falciform process, and anteriorly fusing to a remnant of the lateral lamina of the pterygoid (see ‘Pterygoid’ below). A very small portion of the orbitosphenoid is visible on the right ventral side of the skull, emerging ventral to the posterior lamina of the pterygoid on the medial wall of the orbit and dorsal of the alisphenoid (only visible on the right side of the skull).
Pterygoid
Both pterygoids are incomplete, missing the pterygoid hamulus and almost all of the lateral lamina. In ventral view, the medial lamina is an extremely thin sheet, meeting the vomer/palatine anterior to the nares and curving posterolaterally to form the anterior and lateral walls of the external bony nares, slightly overlying the palatine ( Fig. 3 View Figure 3 ). The posterior lamina rises ventrolaterally as a thin plate, ventrally concave, and forming the posterior wall of the external bony nares. The posterior lamina is bordered posteromedially by the anterior basioccipital crest, separately by a widely open suture. This open suture is unusual among fossil and living odontocetes, and may represent either an ontogenetic feature or diagnostic feature for Arktocara. The posterior lamina is bordered posterolaterally by the alisphenoid.
Platanistoids bear a bony structure on the ventral side of their skulls: a thin, bony lamina that extends from the ventral surface of the hard palate and runs parallel to the posterior lateral lamina of the pterygoid to finally attach to the alisphenoid medial of the squamosal in the ear region. The holotype of Arktocara yakataga is mainly missing this feature, except for a process protruding from the alisphenoid medial of the subtemporal crest. We interpret this feature as an eroded remnant of the lateral lamina of the pterygoid, that is fused to the alisphenoid (there is no detectable suture line between the two elements in Arktocara). For further discussion of the lateral lamina of pterygoid as a platanistoid feature, see discussion in ‘‘Platanistoid systematics.’’
Palatine
Both of the palatine bones are missing large portions, including the palatine surface, maxillary process, palatal crest, and lateral lamina. However, in ventral view a prominent pterygoid sinus fossa for the hamular lobe is formed by the palatine, anterior to the medial pterygoid lamina and posterior to the palatine groove of the maxilla ( Fig. 3 View Figure 3 ). These grooves represent the minimum extent of the palatine extending anteriorly over the maxilla, since the maxillopalatine suture is impossible to detect. A very small fragment of the horizontal portion of the palatine remains immediately ventral to the ventral infraorbital foramen, partially covered by the medial lamina of the pterygoid.
Squamosal
In dorsal view, the short, wide, and rounded zygomatic process of the squamosal points anterolaterally ( Fig. 2 View Figure 2 ). The temporal fossa is floored by the squamosal fossa, forming narrow valley between the supramastoid crest and the squamosal plate. In lateral view, the squamosal plate is a thin sheet, slightly convex laterally, and overlaps the parietal at an indistinct, rounded suture traversing the temporal fossa ( Fig. 4 View Figure 4 ). The zygomatic process is rounded off. The postglenoid process is greatly reduced and missing its ventral edge on the right side, the postglenoid notch is either absent or too greatly reduced to determine, and the external auditory meatus is difficult to determine. In posterior view, the squamosal is widely visible lateral to the exoccipital, and the temporal crest where these later two elements meet is well developed ( Fig. 5 View Figure 5 ). In ventral view, glenoid fossa on the zygomatic process is broad and shallow ( Fig. 2 View Figure 2 ). The incomplete postglenoid process is square in cross section. The contribution of the squamosal to the periotic fossa is wide and shallow, sloping medially from the anterior meatal crest, and bordered medially by the parietal exposure in the periotic fossa. The falciform process is transversely thin and flat, and projects ventromedially from the shallow tympanosquamosal recess. Anteromedial of the falciform process, the anterior margin of the squamosal plate and the anterodorsal margin of the falciform process are extended and join to form an anterior protrusion, which overlies the lateral margin of the alisphenoid. This anterior protrusion is observed in other platanistoids, including Platanista and an undescribed platanistoid USNM 214911 (Fig. S1), where it articulates with the alisphenoid and the lateral lamina of the pterygoid Further study will determine if this feature is present in more platanistoid taxa.
Body size estimate
Total body length (TL) was estimated using the formula created by Pyenson & Sponberg (2011) for calculating body size in stem Platanistoidea (sensu Pyenson & Sponberg, 2011) based on a bizygomatic width (BIZYG):
Log(L) D 0:92
(log(BIZYG)
1:51) C 2:49:
The bizygomatic width of USNM 214930 was measured as 19.1 cm, and using the formula produced a reconstructed body length of 2.28 m. Based on this estimate, Arktocara would have been similar to the adult size of Platanista , which averages a length of 2.4 m and at least 85 kg in weight ( Jefferson, Webber & Pitman, 2008). It is likely that, in life, Arktocara possessed a rostrum that was relatively elongate, based on its near relatives Zarhinocetus and Goedertius; the rostrum of Allodelphis is poorly known, based on several incomplete fragments belonging to the type specimen YPM 13408. In addition, all known allodelphinids have elongated necks, indicated by unfused, proportionally large cervical vertebrae with anteroposteriorly elongated centra, which is in contrast to the general cetacean trend of shortened, hydrodynamic necks ( Buchholtz, 2001). Though the type specimen of Arktocara lacks any postcranial material, it most likely also had an elongated neck given its close relationships to other allodelphinids. Such longirostry and neck elongation may add to Arktocara’s reconstructed total length, and although Pyenson & Sponberg’ s (2011) equations took such allometry into account, we propose that a TL of 2.28 m for Arktocara may be a slight underestimate.
Ontogeny
We assessed skeletal maturity based on longestablished osteological indicators, particularly the fusion of cranial sutures and textural surface of the occipital condyles ( Pyenson & Sponberg, 2011). Most sutures are clearly distinguishable and closed, with some exception of sutures on the ventral side of the skull that appear unsutured. Most pronounced are the open sutures between the dorsal lamina of the pterygoids and the basioccipital on the medial ventral surface ( Fig. 3 View Figure 3 ). It is unclear whether this feature is an ontogenetic trait unique to Arktocara, or whether it is more broadly observed in other allodelphinids (for example, Zarhinocetus). Also, the missing nasals and palatines suggest that their sutures to adjacent skeletal elements were unfused. Pyenson & Sponberg (2011) described the presence of a pitted periosteal surface of the occipital condyles as an indication of immaturity. The preserved occipital condyles of USNM 214830 are smooth, indicated a more advanced ontogenetic age. Based on these combined observations, we suggest that the skull of USNM 214930 belonged to a skeletally mature individual.
Phylogenetic analysis results
The phylogenetic analysis resulted in 430 most parsimonious trees, all with a score of 1960, consistency index of 0.233 and retention index of 0.631. The strict consensus tree, which was created from the 430 trees, shows a similar topology to the equally weighted analysis of Tanaka & Fordyce (2015a). Arktocara is the sister taxon to Allodelphis, nested within a broader clade of Allodelphinidae , which includes Zarhinocetus and Goedertius. This is the first phylogenetic analysis to include these latter two genera, which were not included in Barnes (2006) ’s original matrix. Lambert, Bianucci & Urbina (2014) recovered a monophyletic Allodelphinidae in their 50% majority consensus tree and Lambert, Muizon & Bianucci (2015) resolved a polyphyletic Allodelphinidae , although both analyses only included Zarhinocetus and Allodelphis among their allodelphinid sample of Platanistoidea. Our analysis yields robust support for the monophyly of Allodelphinidae , with higher support values (decay index 5, bootstrap 50) than those recovered for the nodebased clade of Platanistoidea (decay index 1, bootstrap value <50) ( Fig. 10 View Figure 10 ). Like Tanaka & Fordyce (2014) and Tanaka & Fordyce (2015a), we failed to recover a monophyletic Squalodelphinidae (sensu Lambert, Bianucci & Urbina, 2014), yet in contrast, we did find low support for a monophyletic Waipatiidae , an idea proposed by Fordyce (1994), but not explicitly tested until recently. Our analysis is not the first one to recover a clade that includes both species of Waipatia and both species of Otekaikea Tanaka & Fordyce’ s (2015a; 2015b) results are consistent with ours however Tanaka and Fordyce excluded the Otekaikea species from their definition of Waipatiidae . We recommend defining Waipatiidae more inclusively (both species of Waipatia and Otekaikea).
Though it is beyond the scope of this study, the incorporation of molecular data into future analyses of odontocete systematics will help to better resolve the relationships among extant cetaceans, and consequently their fossil relatives. See below for further comment on the implications of these results on the systematics of Platanistoidea.
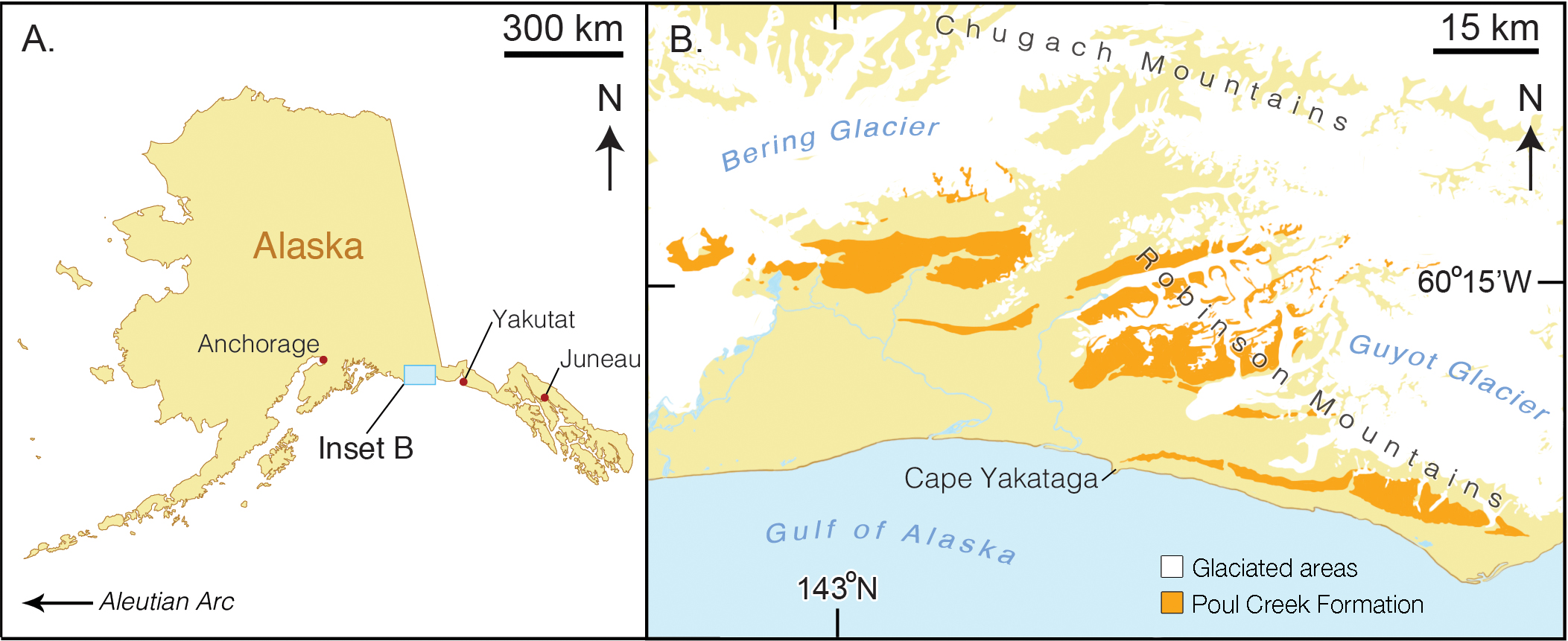

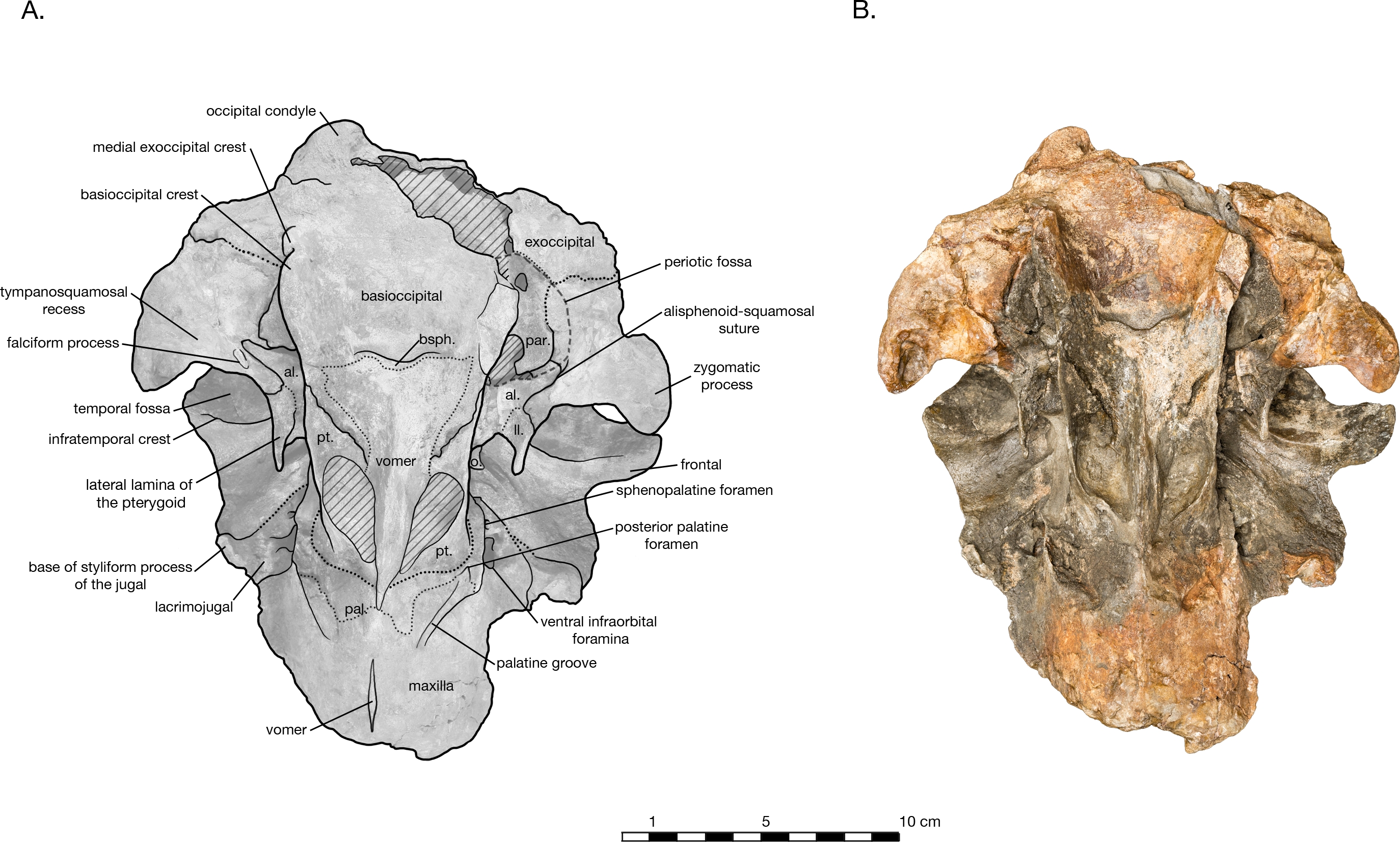
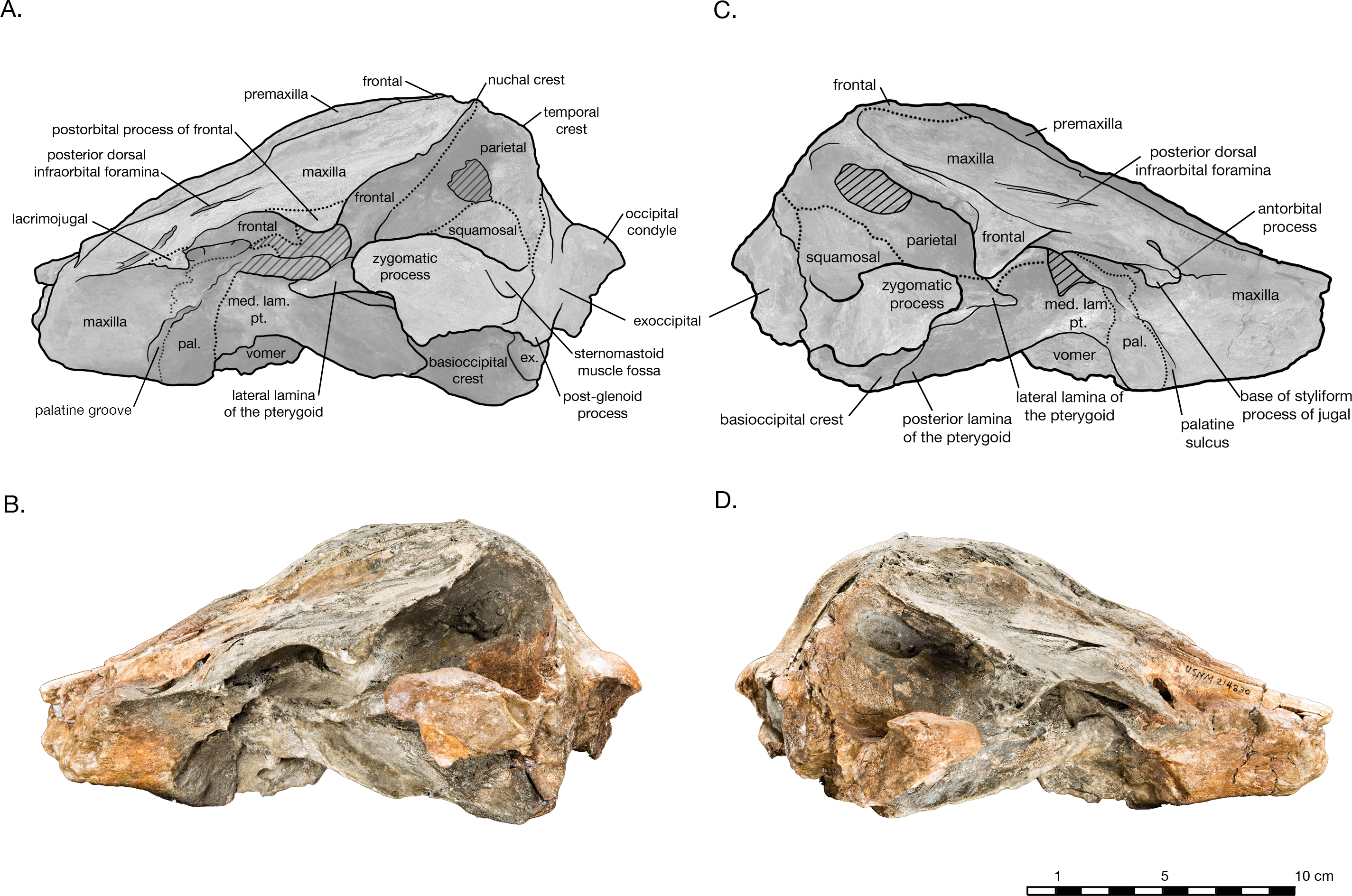
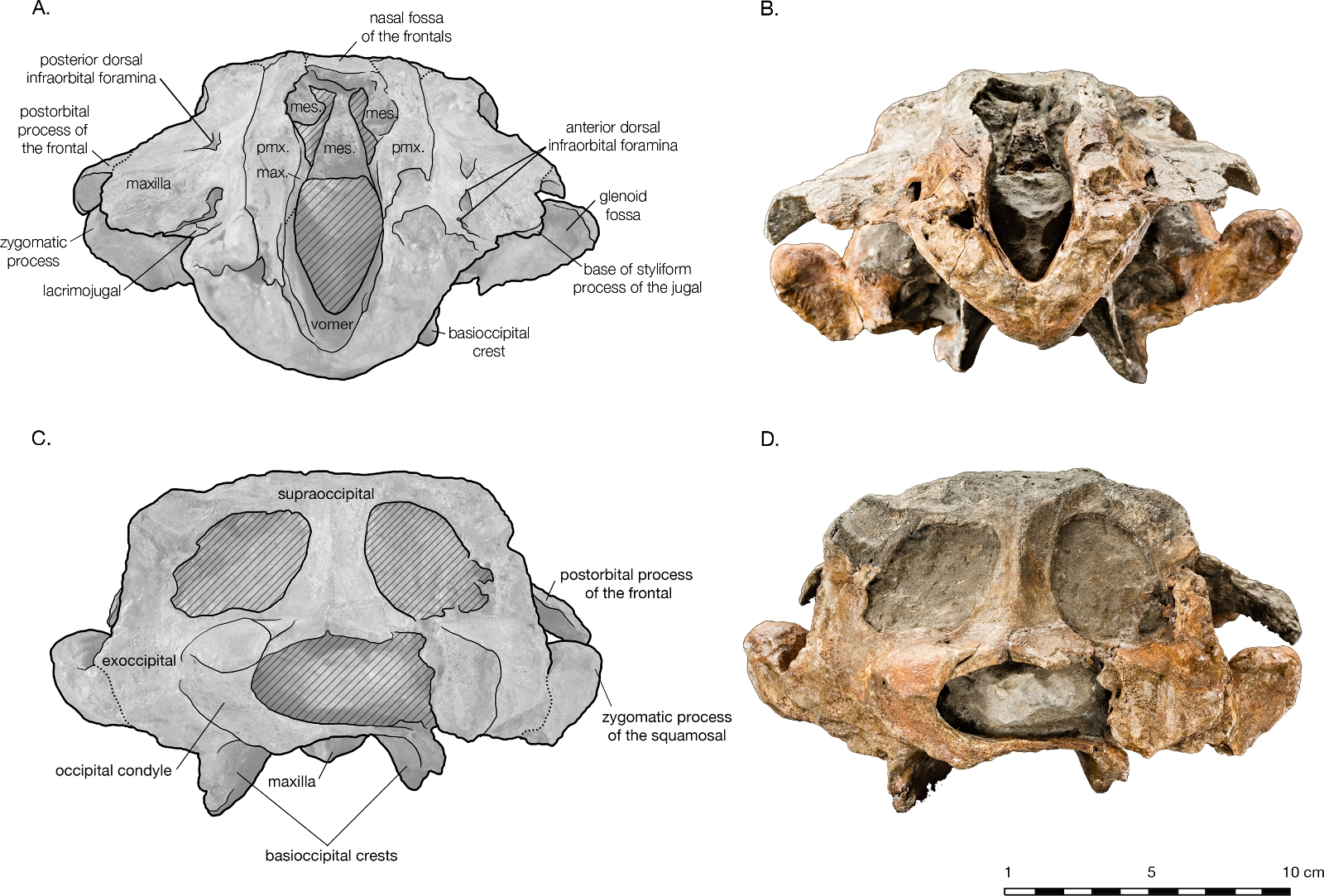


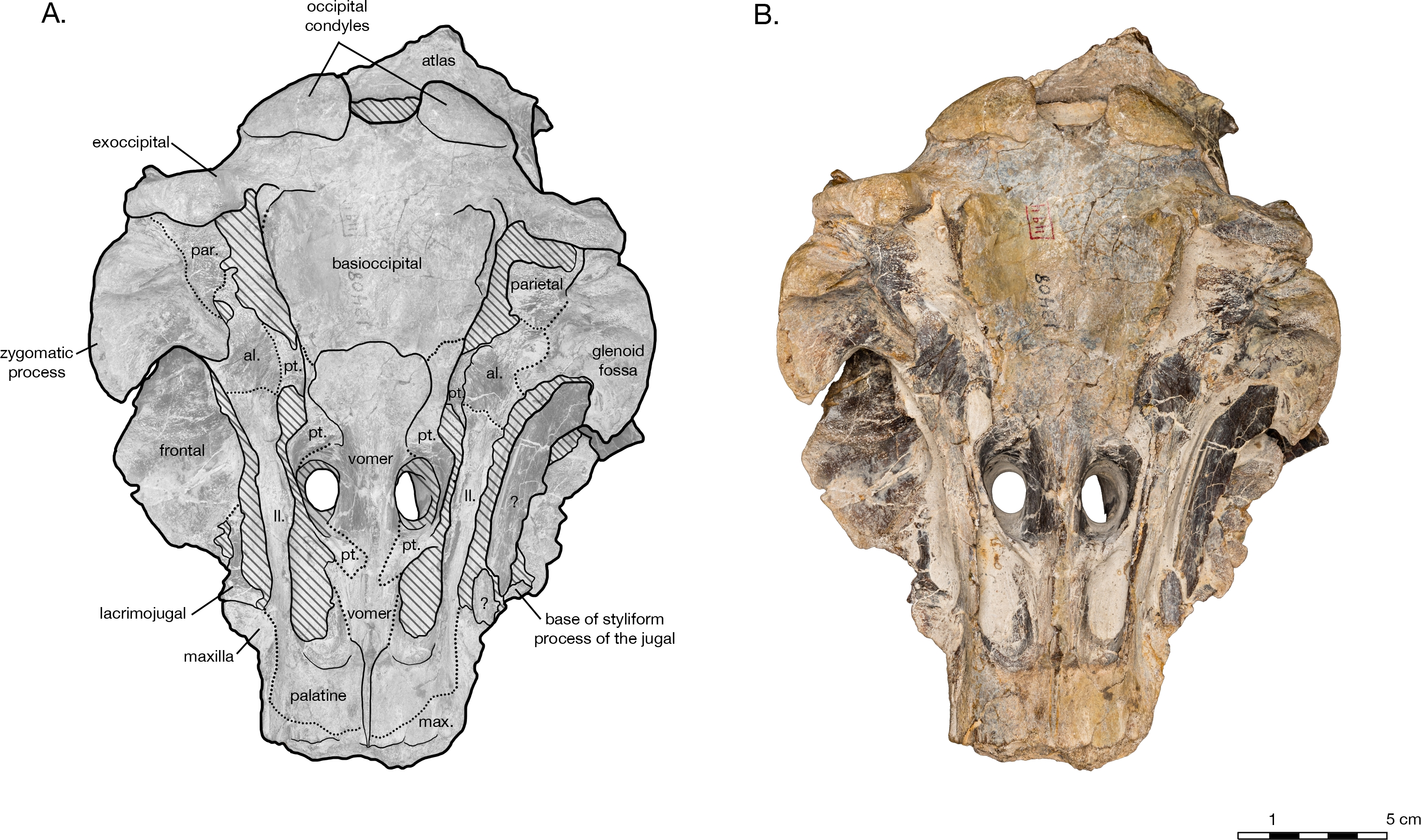


Table 1 Measurements for type specimen Arktocara yakataga (USNM 214830). Measurements of holotype skull of Arktocara yakataga (USNM 214830), in cm (modified after Perrin, 1975 and Tanaka & Fordyce, 2014).
| Dimension | Measurement (in cm) | ||
|---|---|---|---|
| Total preserved length of skull from furthest anterior point to furthest posterior point | 23 | ||
| Cranial length from antorbital notches to occipital condyle | Distance from preserved rostrum tip to external nares (to mesial end of anterior transverse margin of right naris) | 17.5 | 8.5 |
| Distance from preserved rostrum tip to external nares (to mesial end of anterior transverse margin of right naris) | 8.5 | ||
| Distance between upper margin of foramen magnum and nuchal crest | 6.8 | ||
| Height of foramen magnum | 2.9 | ||
| Height of occipital condyle | 4.1 | ||
| Height of temporal fossa | 5.9 | ||
| Height of rostrum at base | 6.9 | ||
| Length of temporal fossa | 5.8 | ||
| Orbit length | 4.9 | ||
| Maximum length of nasal fossa of the frontal | 2.6 | ||
| Length of vertex (nuchal crest to anterior transverse margin of nasal fossa of the frontal) | 4 | ||
| Depth of rostrum at base | 6.5 | ||
| Width of rostrum between antorbital notches | 8.8 | ||
| Width of premaxillae at rostrum base | 5.5 | ||
| Maximum width of premaxillae on cranium | 6.2 | ||
| Width of external bony nares | 3.6 | ||
| Postorbital width of skull | 17.7 | ||
| Bizygomatic width of skull | 19.1 | ||
| Width between temporal crests | 11.1 | ||
| Width of foramen magnum | 5.7 | ||
| Width of occipital condyles | 9.8 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.