
Prolachlanius resinatus (Hagen in Pictet-Baraban et Hagen, 1856 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4688.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:678D7182-DAF4-4AC8-ACC1-1921DA78EF36 |
|
DOI |
https://doi.org/10.5281/zenodo.5944920 |
|
persistent identifier |
https://treatment.plazi.org/id/E1388780-3215-FFD6-A2FC-FD43CD3E4B8A |
|
treatment provided by |
Plazi |
|
scientific name |
Prolachlanius resinatus (Hagen in Pictet-Baraban et Hagen, 1856 ) |
| status |
|
Prolachlanius resinatus (Hagen in Pictet-Baraban et Hagen, 1856) View in CoL
Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6
Type material. Neotype (by present designation) GPIH Typ. Kat. Nr. 4994 (collection of Carsten Gröhn, no. 7322), deposited in GPIH (CeNak). A complete male specimen.
Type locality and horizon. Baltic amber (Yantarnyi [formerly Palmniсken], Kaliningrad Region, Russia). Late Eocene.
Other material. Four specimens from Baltic amber (probably collected near present-day Kaliningrad): GZG. BST. 05240-5858 (possibly a male, but not clearly visible), GZG.BST. 05241-5868 (female), GZG.BST. 05242- 5870 (apparently female); GZG.BST. 05238-5914 (apparently female). All are from the famous amber collection of Geologisch-Paläontologische Institute der Albertus-Universität in Königsberg, a small part of which is deposited in GZG.
Specimen SIZK K-6835 (sex unknown) from the Klesov locality (Pugach quarry) in the Rovno Region, Ukraine (Rovno amber); late Eocene, deposited in SIZK. Syninclusion: Coleoptera ( Corylophidae ?).
Re-description. Head cuneiform in lateral view. Eyes relatively large. Vertex slightly elevated covered with long scarce setae. Terminal segment of maxillary and labial palpi relatively thick, apically abruptly narrowed to form elongate acute portion (forming additional sub-segment?) ( Fig. 3B View FIGURE 3 ). Antennae 35–39-segmented; scape distinctly enlarged and elongated.
Pronotum relatively short, transverse, covered with long, rather scarce setae ( Fig. 3A View FIGURE 3 ). Structure of meso- and metanotum usual for Hemerobiidae .
Foreleg shorter than mid-, hind legs. Femora markedly narrower than tibiae, which are swollen: protibia shorter, less swollen than mesotibia; mesotibia shorter, less swollen than metatibia, which are longest, strongly swollen. Tibial spurs in all legs very short. Tarsi of all legs similarly constructed, with short tarsomeres (basitarsi longest). All legs covered with fine short relatively scarce setae.
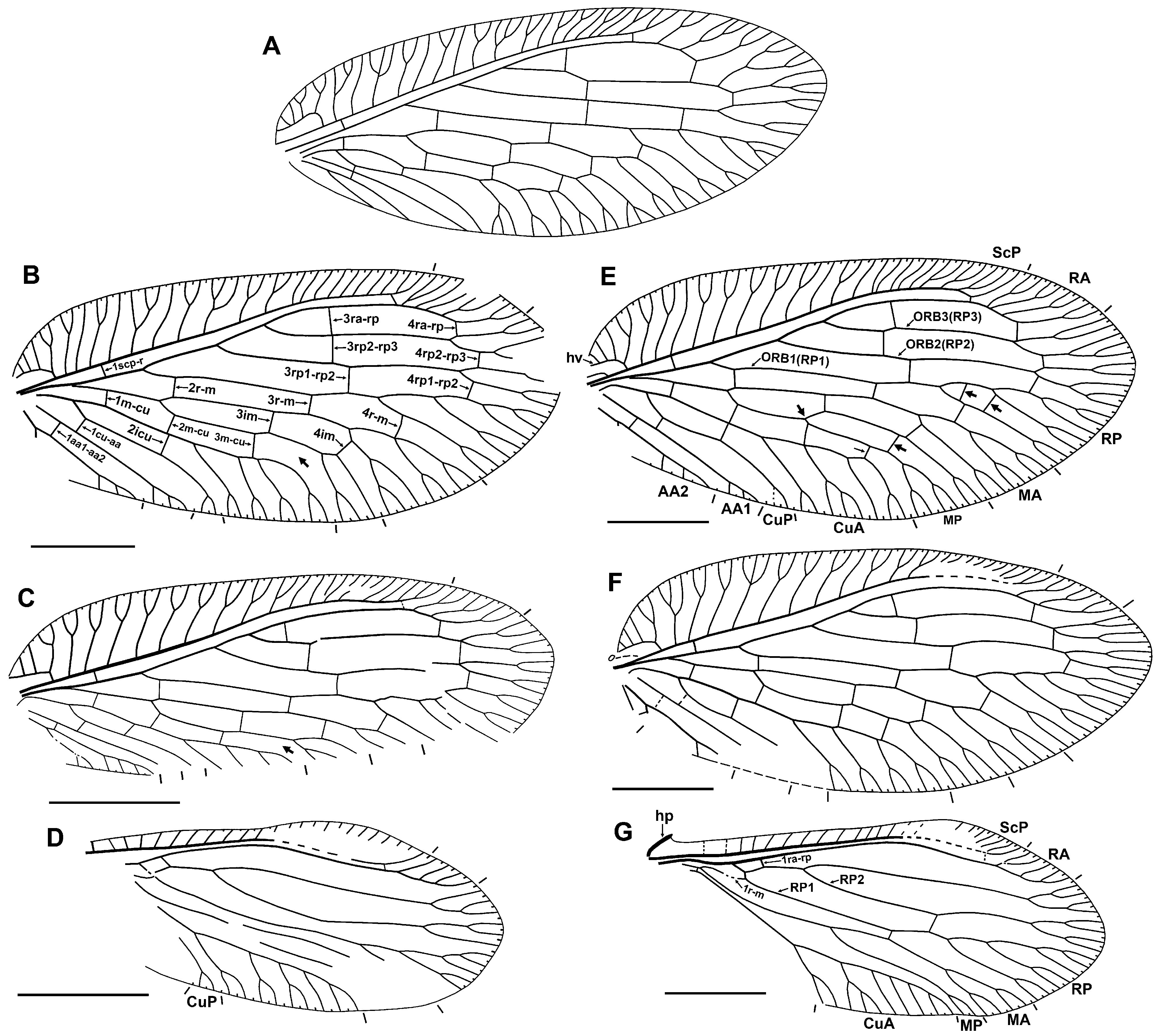
Forewing oval, 4.2–5.6 mm ling, 1.9–2.6 mm wide (length/width ratio 2.15–2.26). Trichosors prominent along entire wing margin. Costal space broad, strongly dilated proximally. Majority of pre-pterostigmal subcostal veinlets forked once or twice; pterostigmal subcostal veinlets mostly simple, some poorly discernible near ScP. Humeral veinlet (i.e., basal-most subcostal veinlet) recurrent, pectinately branched, with four (rarely three, including neotype) branches (usually proximal three simple, distal one forked). True crossveins in costal space absent. Subcostal space moderately wide for entire length, with two crossveins: basal, distal (in Klesov specimen two to three distal crossveins). Posterior trace of RA forked apically, with two to four branches, mostly once forked. Three separate branches of RP originated from R (formally from RA). RP 1 relatively profusely, in general pectinately branched, with two to branches. RP 2, RP 3 dichotomously branched distad fourth gradate series. M forked slightly distad origin of RP 1. MA, MP normally similarly dichotomously forked: deep fork at fourth gradate series, both branches once to twice shallowly forked ( Figs 1A, F View FIGURE 1 , 6C, D View FIGURE 6 ); rarer, these veins have other mode of branching, e.g., MA with more number of branches ( Fig. 5B View FIGURE 5 ), MP more deeply forked at third gradate series ( Fig. 5E View FIGURE 5 ). Anterior trace of CuA forked apically once to twice; pectinately branched, with two to three long branches, most of which once (rarely twice) forked. CuP with only one marginal fork. AA1 shallowly forked once or twice. AA2 deeply forked; anterior branch pectinate, with two to four short simple branches; posterior branch probably pectinate, with one to two very short branches (not clearly discernible in most specimens due to recurved anal region). AA3 simple. Four gradate series of crossveins (series 1 to 4 of Oswald, 1993) posterior to RA. First series consists of four crossveins: 1mcu, 1cu-aa1, 1aa1-aa2, aa2-aa3, arranged similarly as in Sympherobius ; all relatively pale. Second series includes three crossveins: 2r-m, 2m-cu, 2icu; of these, latter two dark brown. Third (‘inner’) series with six crossveins (from 3ra-rp to 3m-cu); 3rp1-rp2 usually shifted distally relative to next crossveins in series; all brown to dark brown. Fourth (‘outer’) series normally complete, with six crossveins (from 4ra-rp to 4m-cu); of these three posterior darker (brown) than anterior three, which are relatively pale. Crossveins absent between branches of CuA. Wing membrane brownish, rarely pale; some crossveins (especially 3ra-rp, 3rp2-rp3, 3rp1-rp2, 2m-cu, 2icu) margined with dark brown, narrowly (e.g., Figs 1A View FIGURE 1 , 4C View FIGURE 4 , 6A, B View FIGURE 6 ) to more broadly (e.g., Figs 4A, D View FIGURE 4 ); posterior part of pterostigma with dark brown shading, more or less broad (e.g., Fig. 4B View FIGURE 4 ) to narrow (e.g., Figs 1 View FIGURE 1 , 4C, D View FIGURE 4 , 6A, B View FIGURE 6 ).
Hind wing ( Figs 5D, G View FIGURE 5 , 6E View FIGURE 6 ) ca. 3.3–3.4 mm long, 1.4 mm wide (neotype). Humeral plate well developed. Trichosors prominent along entire margin. Costal space narrow, dilated in pterostigmal area (in neotype and GZG. BST.05240-5858 strongly dilated). Subcostal veinlets simple, these in pterostigmal area poorly discernibly near ScP. Subcostal space narrow, with one distal crossvein (in Klesov specimen two distal crossveins). RA with two to three distal veinlets. Crossvein 1ra-rp relatively short to moderately long, perpendicular to RP. RP with two branches. Anterior trace of RP and RP 2 dichotomously branched, with two to three forks each. RP 1 pectinately branched, with two to three branches. Basal crossveins 1r-m sinuous, connecting M with RP 1. Two crossveins in inner gradate series, none in other. M forked distad origin of most proximal branch of CuA and of RP 2. Anterior trace of CuA forked distally, with four to five pectinate branches (of these, two to three branches forked once or twice (rarely), other simple) (based on neotype and specimen GZG.BST.05240-5858). CuP present, forked terminally. Anal veins not clearly discernible in any specimens, as region hidden by forewings or poorly preserved. All veins pale brownish; wing membrane pale brownish to nearly hyaline; pterostigmal area more fuscous, partially blackish in neotype ( Fig. 1B View FIGURE 1 ).
Abdominal details poorly discernible in most specimens, especially their terminal segments.
Male (neotype). Sixth, seventh tergites, sternites not specialized (tergites little extended laterally, sternites more strongly so). Eighth tergite relatively short, stronger extending laterally than previous tergites. Eighth sternite very narrow, extending laterally like seventh sternite. Ninth tergite narrow, strong extending laterally, ventrally with acute apices (clearly visibly on right side ( Fig. 2B View FIGURE 2 ), but this portion on left side poorly preserved ( Fig. 3D View FIGURE 3 )). Ninth sternite relatively small, short, shifted far proximally; surface distad it smooth, lacking any trichiation, even microtrichia (this is possibly visible apical part of parabaculum (=gonocoxites + gonapophyses 10 of Aspöck & Aspöck 2008)). Ectoprocts elongate, directed ventrally; its dorsal part dilated narrowing ventrad; apically distinctly dilated, inwardly curved, furnished with ten spines and strong setae ( Fig. 3C View FIGURE 3 ). Callus cerci rounded (oval), number of trichobothria hard to count (probably>20). All segment covered with macro- and microtichiae; setae especially long, dense ventrally on sixth to eighth sternites. Presumable mediuncus (=gonostyli 9 of Aspöck & Aspöck 2008) well developed extending far from ectoprocts; appears unpaired oval structure (in lateral view) with acute apex, which is bent ventrally.
Female. Nine tergite strongly dilated ventrally. Ectoproct large, more or less rounded. Gonostylus 9 rather long. Other details not clearly discernible.
Remarks. Prolachlanius resinatus was described from five specimens (see Pictet-Baraban & Hagen 1856), but Krüger (1923) found that only one belongs to this species, which he designated as ‘Arttype’ (i.e., species’ type, or the holotype). Five other specimens labeled “ resinatus ” belong to other orders (one Lepidoptera and four Trichoptera), these are probably not from the type series. He assumed that the other four specimens of the type series belong to Proberotha prisca Krüger, 1923 , because one of these was labelled “ Hemerobius ”. It is interesting to note that Pictet-Baraban & Hagen (1856) and Krüger (1923) possibly examined two different specimens of Prolachlanius resinatus . Hagen wrote that there are six crossveins in the fourth gradate series, and the figure verifies it ( Pictet-Baraban & Hagen 1856: Pl. 8, fig. 17). In contrast, Krüger mentioned only five of these crossveins, similar to the condition seen in the neotype and specimen GZG.BST.05241/5868 ( Fig. 5B, C View FIGURE 5 ).
The holotype has not been located in German or Polish museums, and appears to be lost. Here, we designate a male with a well-preserved abdomen with clearly visible terminal segments as its neotype to avoid probable future taxonomic misunderstandings. The neotype and the lost holotype have very similar forewing venation; they are undoubtedly conspecific.
This species is apparently sexually dimorphic in the structure of the hind wing pterostigmal area. At least, this is evidently dilated and strongly fuscous in the male (neotype) so that the costal margin in pterostigmal area is convex ( Fig. 5D View FIGURE 5 ), while this area in the female (at least in GZG.BST. 05241-5868: Fig. 4D View FIGURE 4 ) is not dilated and only slightly fuscous, and the margin is not convex. If this hypothesis is correct, the Rovno amber specimen is a female.
Also, the difference in relative widths of the forewing of this species is probably at least partly explained by sexual dimorphism. In some species of other genera of Hemerobiidae , the forewing in the female is significantly narrower than that of males ( Makarkin & Kholin 1995).
Wing venation and maculation are variable in Prolachlanius resinatus . Specimen GZG.BST.05240/5858 has notably atypical maculation for the species (see Fig. 4A View FIGURE 4 ). Some specimens have almost hyaline wings, although with distinct maculation, but a few have a darker, nearly single-colored membrane lacking distinct maculation ( Fig. 7 View FIGURE 7 ). Of these, specimen T079 is noteworthy by the absence of crossveins 4r-m, 4im and 4m-cu in the left forewing, i.e., a condition characteristic of the genus Sympherobius (also see below). Its forewing is 4.6 mm long, i.e., slightly shorter than in more typical specimens. It is hard to decide if specimen T079 represents an atypical Prolachlanius resinatus or a new species.
Some venational aberrations should be mentioned. In one specimen from a private collection with typical maculation and otherwise typical venation, CuA has only one branch. In specimen GZG.BST.05238/5914 there are a number of anomalies, including MP being deeply forked at the third gradate series ( Figs 4B View FIGURE 4 , 5E View FIGURE 5 ).
The forewings of the Klesov specimen (Rovno amber) have generally very similar venation to those of most specimens from Baltic amber, but differ from these by [1] more reduced dark bordering of crossveins 3ra-rp, 3rp2-rp3 and 3rp1-rp2, [2] the loss of crossvein 4rp1-rp2, and [2] a greater number of distal subcostal crossveins ( Fig. 6 View FIGURE 6 ).
[1] The Klesov specimen is probably more similar to the holotype than it is to some other Baltic amber specimens by this character state; at least, Pictet-Baraban and Hagen (1856) and Krüger (1923) do not mention the crossveins having dark borders. Hagen wrote only that “wings are hyaline and some gradate crossveins are dark, in [the] pterostigma [there is a] brown-shaded spot” [Die Flügel sind wasserklar und einzelne Stufenadern dunkel, im Pterostigma ein brauner Schattenfleck] (p. 88), i.e., the holotype maculation is very similar to that of the Klesov specimen, but the membrane of both fore- and hind wings in the Klesov specimen and most Baltic amber specimens (including the neotype) are clearly slightly brownish.
[2] The loss of crossvein 4rp1-rp2 is most probably an anomaly as the loss of almost any crossvein may occur in individual extant hemerobiids.
[3] The presence of two to three distal subcostal crossveins in the Klesov specimen is noteworthy, although this is not a specific character state in this family. The number of these crossveins vary strongly within species of extant Hemerobiidae , e.g., there are one to six distal crossveins in Micromus angulatus ( Stephens, 1836) ; two to eight in Micromus dissimilis ( Nakahara, 1915) ; and two to three in Hemerobius tristriatus Kuwayama, 1954 (VM, pers. obs.). However, only one distal subcostal crossvein is present in all extant Sympherobiinae . At least, the presence of two or more is unknown to occur in any extant specimens of Sympherobius , even as an anomaly.
Therefore, character state [3] may have some taxonomic value. However, to evaluate how much value, it is necessary to study more specimens from Rovno amber.






| MP |
Mohonk Preserve, Inc. |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.