
Nesozetidae, Hammer, 1971
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4358.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:2A99B4C9-5AFD-4C4B-B3AB-E80AAFC555EA |
|
DOI |
https://doi.org/10.5281/zenodo.6044424 |
|
persistent identifier |
https://treatment.plazi.org/id/E16287AA-FFA5-901B-FF0A-F8F5FBC1F843 |
|
treatment provided by |
Plazi |
|
scientific name |
Nesozetidae |
| status |
|
Morphology of Nesozetidae View in CoL
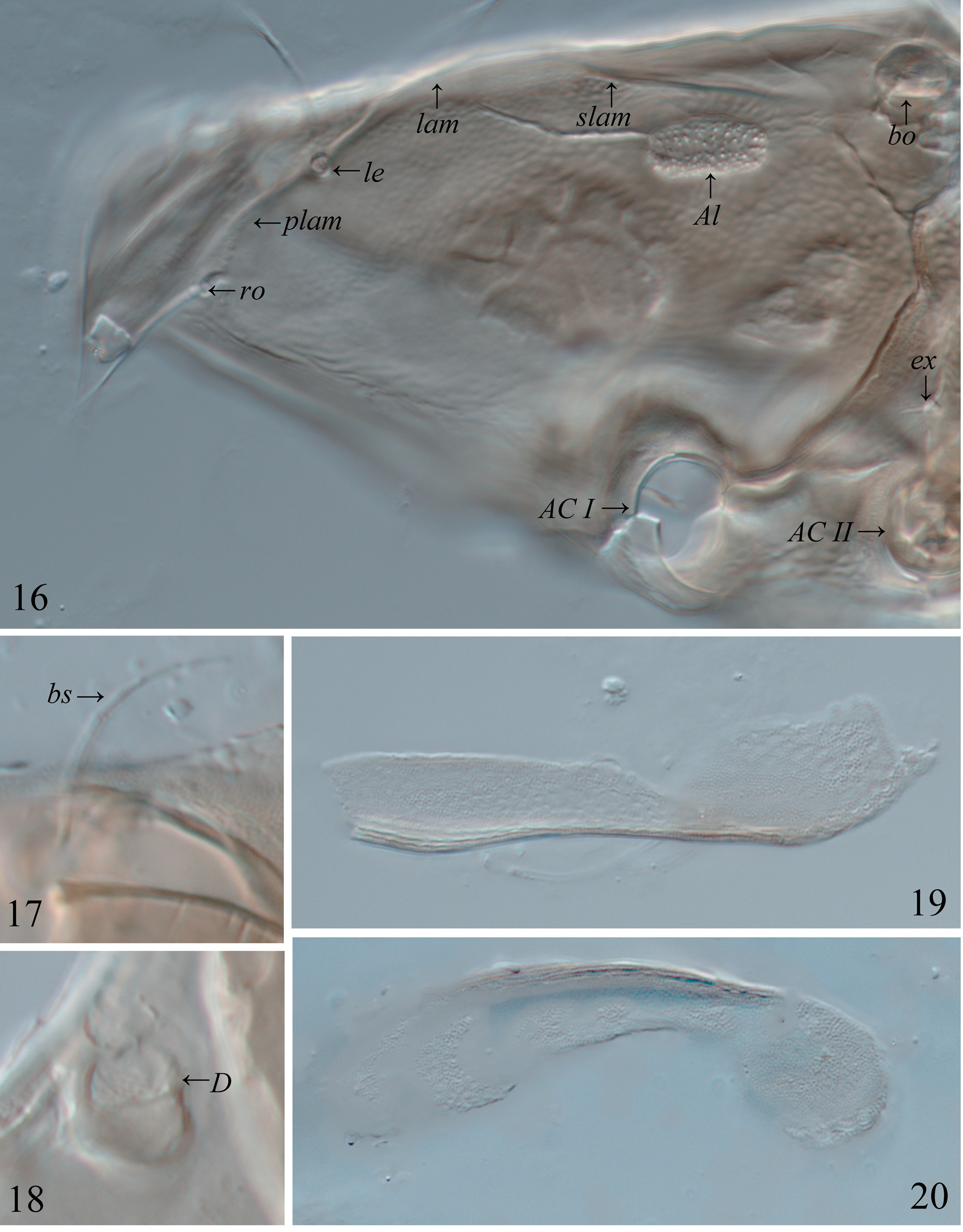
Membranes. DeVelopment of hyaline membranes in adult Nesozetidae is unique in Oribatida and is an autapomorphy for Nesozetidae . These membranes are cerotegumental extensions of the body, and they are somewhat inflexible and easily separated from the body. Similar to cerotegument in some other Brachypylina , e.g., Eremaeozetidae ( Schatz et al. 2006) , they can retain the pattern of the underlying integument ( Figs 19, 20 View FIGURES16–20 ). They are not formed of sclerotized integument and yet retain their structure and are consistently found in the same position on all adults examined, that is associated with the region of the acetabula where the tracheal system opens ( Norton et al. 1997).
The consistency of these cerotegumental membranes in species of Nesozetes , their association with acetabula and the habitat of these species in wet moss suggest that these membranes could be part of a plastron system. A plastron is defined as an alternatiVe respiration system absorbing oxygen from the surrounding water Via a thin layer of air trapped by hydrophobic hairs or other cuticular projections ( Fielden et al. 2011). In most adult Brachypylina plastron function depends on the hydrophobic properties of modified cerotegument, which may or may not be in contact with the tracheal system ( Pugh et al. 1987, Pfingstl & Krisper 2014). For example, the cerotegument of Ameronothrus marinus ( Banks, 1896) contains approx. 20.9% airspace, that of Ameronothrus maculatus ( Michael, 1882) 60.4%, and that of freshwater Hydrozetes Berlese, 1902 66% ( Pugh et al. 1987). Details of the cerotegumental plastron in both adult and immature Fortuyniidae and Selenoribatidae were beautifully illustrated with SEM by Pfingstl & Krisper (2014). Unfortunately, we do not haVe that kind of detail for the species of Nesozetes but future studies of Nesozetes should include obserVation of liVing specimens under flooded conditions to solVe the question of a plastron function.
Leg setation. Adult leg setation is only known for Nesozetes membranus sp. nov. It is a reduced setation and we compare it with setation in other Oripodoidea. Leg setation and its ontogeny haVe been described for few species in the Oripodoidea, and this has limited analyses of relationships within the superfamily. The setation of Nesozetes membranus is only known for the adult, but is reduced in comparison with that of described Oripodoidea, as noted below:
(a) Tarsus I. Setae (a) and tc’ are absent from tarsus I, a combination unique in known Oripodoidea. Setae (a) are absent from this segment in species of Symbioribates Aoki, 1966 (Symbioribatidae) (Karasawa & Behan- Pelletier 2007). In addition, setal pair (pl) and seta pv” do not deVelop in Symbioribates ( Aoki 1966) .
(b) Tarsus II. Setae (a) and tc’ are absent from tarsus II, a combination unique in known Oripodoidea. Setal pairs (a) and (pv) are absent from this segment in Symbioribates ( Karasawa & Behan-Pelletier 2007) . Setae (pv) are also absent from tarsus II of Pirnodus detectidens Grandjean, 1956 , and Truncopes optatus Grandjean, 1956 (Oripodidae) and Dometorina plantivaga ( Berlese, 1896) (Scheloribatidae) , and from Reductobates perangustus ErmiloV & Minor, 2016 ( Haplozetidae ). As Grandjean (1950) noted, loss of fundamental setae such as (a) and (pv) is unusual in Oribatida .
Solenidion ω2 is absent from this segment, as it is in Parapirnodus coniferinus Behan-Pelletier, Clayton & Humble, 2002 , though it is present in the congener Parapirnodus prosopis Martinez, Fernandez & Monetti, 1996 . We retain Parapirnodus coniferinus and Parapirnodus hexaporosus Behan-Pelletier, Clayton & Humble, 2002 in the scheloribatid genus Parapirnodus (Scheloribatidae) rather than in Behanpseudoppia Subías, 2017 ( Pseudoppiidae ) (Subías 2017) pending reVision of the cluster of oripodoid families: Parapirnodidae , Pseudoppiidae , Caloppiidae and Hemileiidae recognized by Subías (2004, 2017), but those genera are all included in Scheloribatidae by Schatz et al. (2011) or Norton & ErmiloV (2014). The solenidion is also absent in Reductobates perangustus .
(c) Tarsus III. Setae (a) and (it) are absent from tarsus III, a combination unique in known Oripodoidea. Setae (it) are also absent in species of Symbioribates . In contrast, setae (pv) are absent from Reductobates perangustus . Iteral setae are also absent from tarsi I to IV in certain species of the families Zetorchestidae , Hermanniellidae and Plasmobatidae ( Grandjean 1961, 1964) and Adhaesozetidae ( Walter & Behan-Pelletier 1993) . As none of these latter families is a member of the Oripodoidea, the iteral setae are considered to haVe been lost independently from tarsi III in Nesozetidae .
(d) Tarsus IV. Setae (a) are absent from tarsus IV. Setae (a) are also absent in R. perangustus .
(e) Tibiae. Tibiae I: Setae v” absent. Tibiae II: setae l” and v” absent. Tibiae III: setae l’ absent. Tibiae IV: setae l’ absent. The tibial setation of 3(2)–2(1)–2(1)–2(1) is the most reduced known among Oripodoidea. The usual setation for this segment in Oripodoidea (and Brachypylina ) is 4(2)–4(1)–3(1)–3(1), e.g., Grandjean (1956). The tibial setation for Symbioribates is almost as reduced, but it retains seta l’ on tibia III (Karasawa & Behan- Pelletier 2007).
(f) Genua III: setae absent. Setae are also absent from this segment in P. coniferinus ( Behan-Pelletier et al. 2002) .
(g) Trochanters I–IV: setae absent. This is unique among known Oripodoidea.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.