
Eusarsiella hanguk, Karanovic, Ivana & Soh, Ho-Young, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3947.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:E0151A24-761D-4465-BB71-DAC4FA5B9550 |
|
DOI |
https://doi.org/10.5281/zenodo.5658721 |
|
persistent identifier |
https://treatment.plazi.org/id/E2798D5D-4A70-FFC4-C8D7-5341280DFBC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Eusarsiella hanguk |
| status |
sp. nov. |
Eusarsiella hanguk View in CoL sp. nov.
( Figures 1 View FIGURE 1 B, 4–7)
Etymology. The species name is the Korean word “ Hanguk ”, meaning Korea.
Material examined. Holotype female dissected on one slide, shell on SEM stub ( NIBR IV 0000287232); two paratype females dissected on one slide each, shell in ethyl alcohol ( NIBR IV 0 0 0 0 287233, and NIBR IV 0000287234), 10 paratypes (two females and eight juveniles) in ethyl alcohol ( NIBR IV 0000287235), and four paratypes used for DNA sequencing from (type locality) South Korea, East China Sea, Jeju Strait, Chuja Island, 33°43’37.954”N 126°21’36.641”E, 29 November 2012, collector H. Y. Soh.
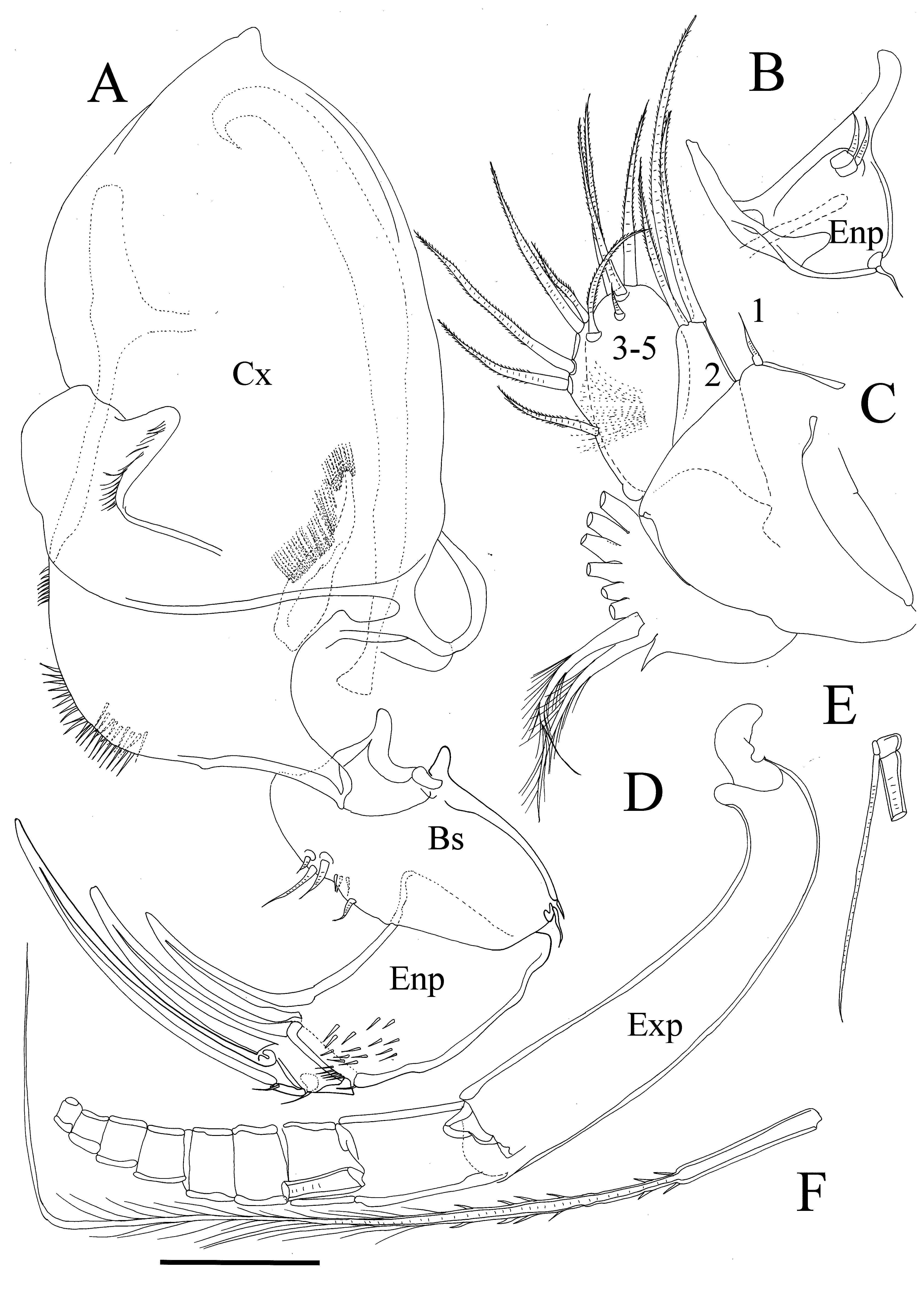
Description. Female. Shell ( Figures 1 View FIGURE 1 B, 4A, B) without ridges, only with wide, shallow pits (fossae). Caudal process not very long and distally pointed. Postero-dorsal bulge clear and rounded, and in lateral view not overpassing dorsal margin. Dorsal and ventral edges of bulge clearly rounded. Dorsal margin of carapace almost evenly rounded. Ventral margin rounded. Long individual bristles distributed on lateral surface of valves, some short and thin, some longer with broad bases. Long pore canals lead from posterior process to inside of valves, shorter ones present along entire anterior and ventral margin ( Fig. 4 View FIGURE 4 B, D). Anterior infold without bristles. Infold of caudal process with only three short bristles and two short ventral ones, and dorsal to caudal process two additional setal bristles present ( Fig. 4 View FIGURE 4 D). Shell L (including caudal process)= 1.45 mm.
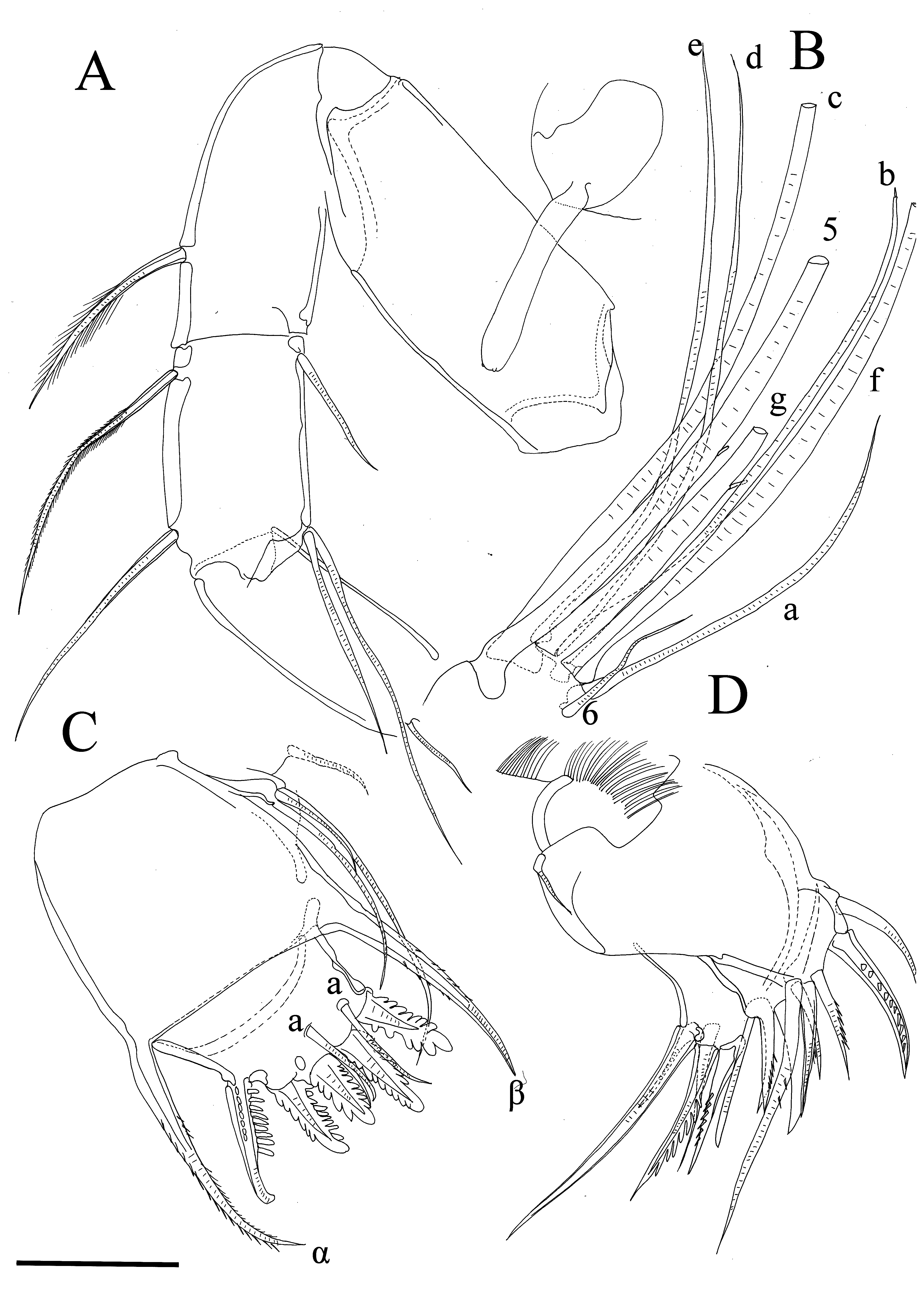
A1 ( Figure 5 View FIGURE 5 A, B). First segment long and bare. Second segment with no dorsal spines and annulated, pappose dorsal bristle reaching 1/3 of following segment. Third and fourth segment fused; third with annulated, pappose dorsal bristle reaching 1/3 of following segment and one shorter and bare ventral bristle (not exceeding distal margin of 3+4 segment). Fourth segment with bare, annulated dorsal bristle not reaching distal end of following segment, and two bare, annulated bristles ventrally (one only reaching distal margin of following segment, other almost 1.5? times longer). Fifth and sixth segments fused. Bristle of fifth segment long, annulated and bare; seta on sixth segment short, also bare. Seventh segment: a-bristle not so short (about three times longer than sixth segment’s bristle); b-bristle slender, with one sensory filament, and about 1/3 longer than a-bristle, this bristle with pointed, sensory-like tip; c-bristle as long as bristle of fifth segment. Eighth segment: d- and e-bristles equally long, bare and shorter than c-bristle; f- and g-bristles slightly shorter than fifth segment bristle, both annulated and gbristle with one sensory filament. All bristles except a-, b-, e- and d- with blunt tips.
Bellonci Organ ( Figure 5 View FIGURE 5 A). Cylindrical, non segmented and short.
Eyes. Both medial and lateral eyes present. Medial eye with scattered pigment, lateral eye with four ommatidia.
A2 ( Figure 6 View FIGURE 6 B, D–F). Prp bare. Enp with two short basal bristles (both annulated and bare), and one very short, bare bristle situated on terminal node. Exp: first segment with small terminal medial recurved tubular bristle; bristles on segments 2–8 with thick spines proximally and with swimming setae distally; ninth segment with two bristles (one short, bare and other longer with swimming setae).
Md ( Figure 6 View FIGURE 6 A). Cx endite elongated and with long spine-like setae, Cx with short slender spines near ventral margin. Bs: ventral margin with seven short bristles, dorsal margin bare with exception of two short marginal slender bristles. First Enp segment with short spines and with one stout claw distally; second Enp segment with one minute spine on dorsal margin, and strong claw ventrally; third Enp segment with stout terminal claw and small dorsal spine.
Mxl ( Figure 5 View FIGURE 5 C, D). Cx with long, bare dorsal bristle. Endite I with three pectinate ringed claws and two ringed bristles; endite II with three pectinate claws and one ringed bristle; endite III with four pectinate claws and two ringed bristles. Exp with three bristles: two long and one short. First Enp segment with spinous alpha- and betabristles. Second Enp segment with five pectinate claws, two a- and one c-bristle (the latter not illustrated).
L5 ( Figure 6 View FIGURE 6 C). Lobe one with one short bristle; lobe two with three bristles (one longer than other two; all armed with tiny setulae); lobes 3–5 fused and with total of 10 bristles, all annulated and with setulae, except most dorsal short bristle. Entire appendage hirsute.
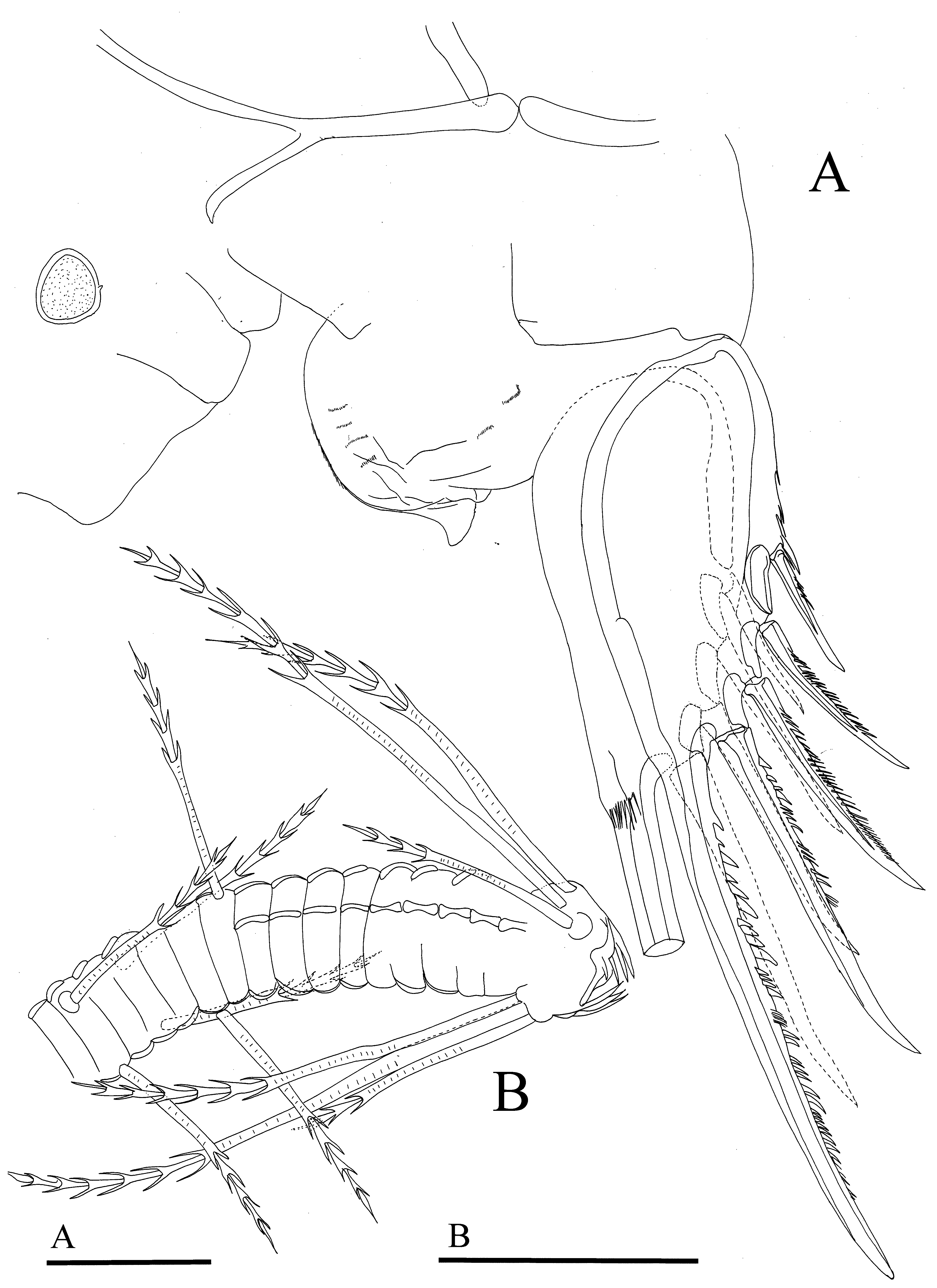
L6 ( Figure 4 View FIGURE 4 C). Single endite with three bristles. Terminal segment with 14 annulated bristles, each with short setulae, and almost all subequally long. Fourteen bristles followed by space and two long plumose bristles. Entire appendage covered with fine, short setulae.
L7 ( Figure 7 View FIGURE 7 B). Proximal group with three bristles on each side, each with four bells. Terminal group with six bristles (three on each side), each with three to six bells. Terminus with opposing combs, each with about five alate teeth.
UL ( Figure 7 View FIGURE 7 A). Each lamella with five claws. Only claw one non-articulated; claws with long and short teeth along posterior edge. Right lamella with group of long, slender spines dorsal to first claw, left lamella with one strong spine following terminal claw. Each lamella with few spines along margin following claws. "Y"-sclerite typical distal branch of the family.
Remarks and affinities. Based on the shape and ornamentation of the shell there is only a handful of species with which E. hanguk may be related to. Namely, the majority of Eusarsiella species have different shell morphologies and they are either smooth, or have ridges, lateral projections, nodules, different shapes of the postero-dorsal extension, different lengths of the caudal process, etc. The following eight species (in alphabetical order) have a very similar carapace shape and ornamentation to the new Korean species: E. asciformis ( Hall, 1987) , E. fallomagna Kornicker, 1994 , E. falx Kornicker, 1992 , E. magna Poulsen, 1965 , E. microthrix ( Chavtur, 1983) , E. reticulata ( Hall, 1987) , E. springthorpei ( Hall, 1987) , and E. tampa Kornicker & Grabe, 2000 .
Three species described from the Great Barrier Reef, Eusarsiella asciformis , E. reticulata , and E. springthorpei , have a very similar carapace shape to E. hanguk . The first species differs from E. hanguk by a slightly finer ornamentation, the presence of three (instead of two) bristles ventrally on the fourth segment of A1, and by the presence of three and one (instead of three and three) bristles proximally on L7. The line drawings of E. asciformis’ carapace (see Fig. 11 View FIGURE 11 A in Hall 1987) show a very similar caudal process as in E. hanguk , while the SEM photos (see Fig. 13 View FIGURE 13 A in Hall 1987) show a much shorter, almost non-existent process. It remains to be tested if this is a variability or Hall (1987) was dealing with two species. Eusarsiella reticulata also has a slightly shorter caudal process. The main difference between this and the new species is in the appearance of the claws on the Md. They are much stronger and thicker than in E. hanguk . There are also other differences, such as the chaetotaxy of the L7. Finally, E. springthorpei differs from the new species by a slightly higher postero-dorsal extension, three setae ventrally on the fourth segment of A1, and a different chaetotaxy of the L7 (with four and five proximal bristles).
Eusarsiella fallomagna was described from New South Wales ( Australia) ( Kornicker 1994), and has a very similar carapace shape, ornamentation, and postero-dorsal extension like E. hanguk but it has more bristles on the caudal infold, three setae ventrally on the fourth segment of A1, and five or six bristles proximally on the L7.
A species from Mozambique ( Kornicker 1992), E. falx , has a similar postero-dorsal extension and similar basic ornamentation, but in addition to fossae its surface is densely covered with small spines. There are some other differences between it and E. hanguk and they mostly refer to the chaetotaxy of L5 to L7.
Eusarsiella magna was described from southeast Australia ( Poulsen 1965) and has almost identical morphology of the carapace with E. hanguk , and the two species practically only differ in the presence of three (instead of two) bristles on the fourth segment of A1 and more (seven or eight) bristles proximally on the L7.
Eusarsiella microthrix , described from Vietnam ( Chavtur 1983), seems to have a similar carapace shape like the new species, but according to the drawings provided in the original description it also appears that the shell was decalcified and slightly deformed, and there is an apparent presence of some short ridge-like structures which may be a result of shell deformation. However, the new species can easily be distinguished from E. microthrix by a different length ratio between the claws on the UL (in the new species the claws are much more slender), and the presence of three and three (instead of two and two) bristles proximally on the L7.
Finally, E. tampa, described from the Gulf of Mexico ( Kornicker & Grabe 2000), has more bristles on the infold of the caudal process, a shorter a-bristle on the A1, only two (instead of three) bristles on the second endite of the L 5 and two and one bristle proximally on the L 7.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.