
Epsilonema rugatum, LORENZEN, 1973
|
publication ID |
https://doi.org/ 10.1111/zoj.12324 |
|
publication LSID |
lsid:zoobank.org:pub:175CF665-173D-40DE-AA2F-8417F520AE22 |
|
persistent identifier |
https://treatment.plazi.org/id/E34287C9-FF93-DB76-821A-1D324890FBD3 |
|
treatment provided by |
Marcus |
|
scientific name |
Epsilonema rugatum |
| status |
comb. nov. |
EPSILONEMA RUGATUM LORENZEN, 1973 , COMB. NOV.
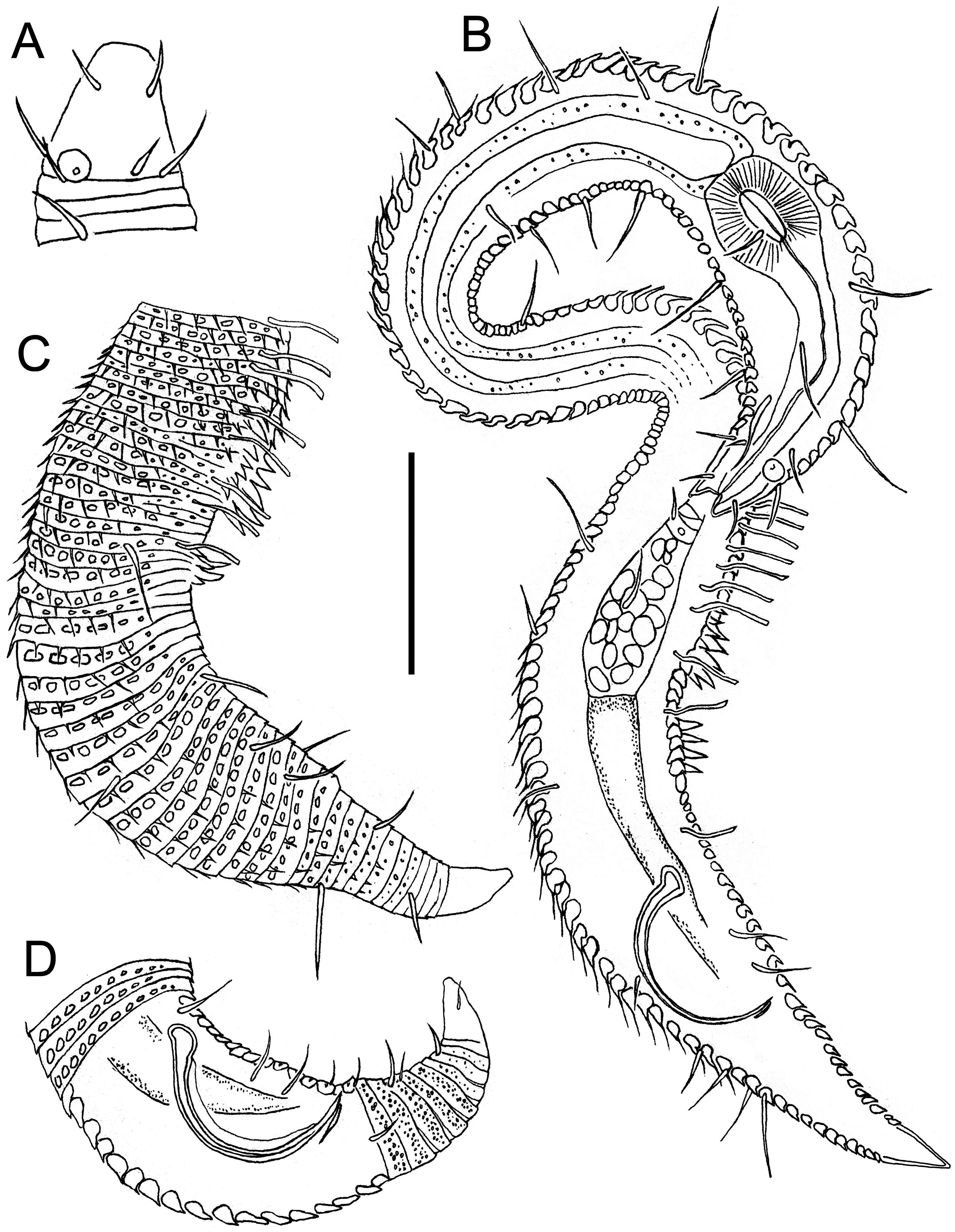
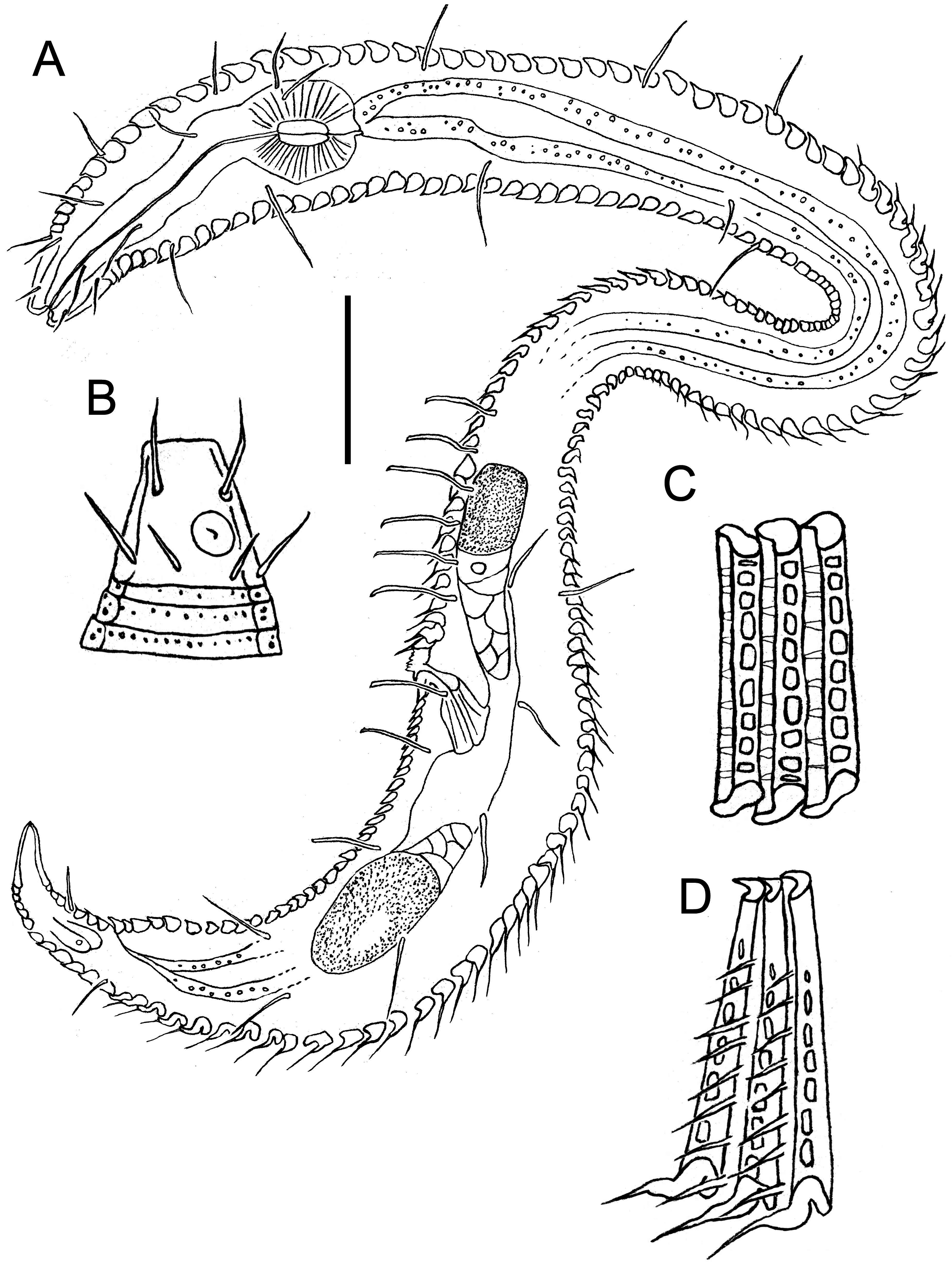
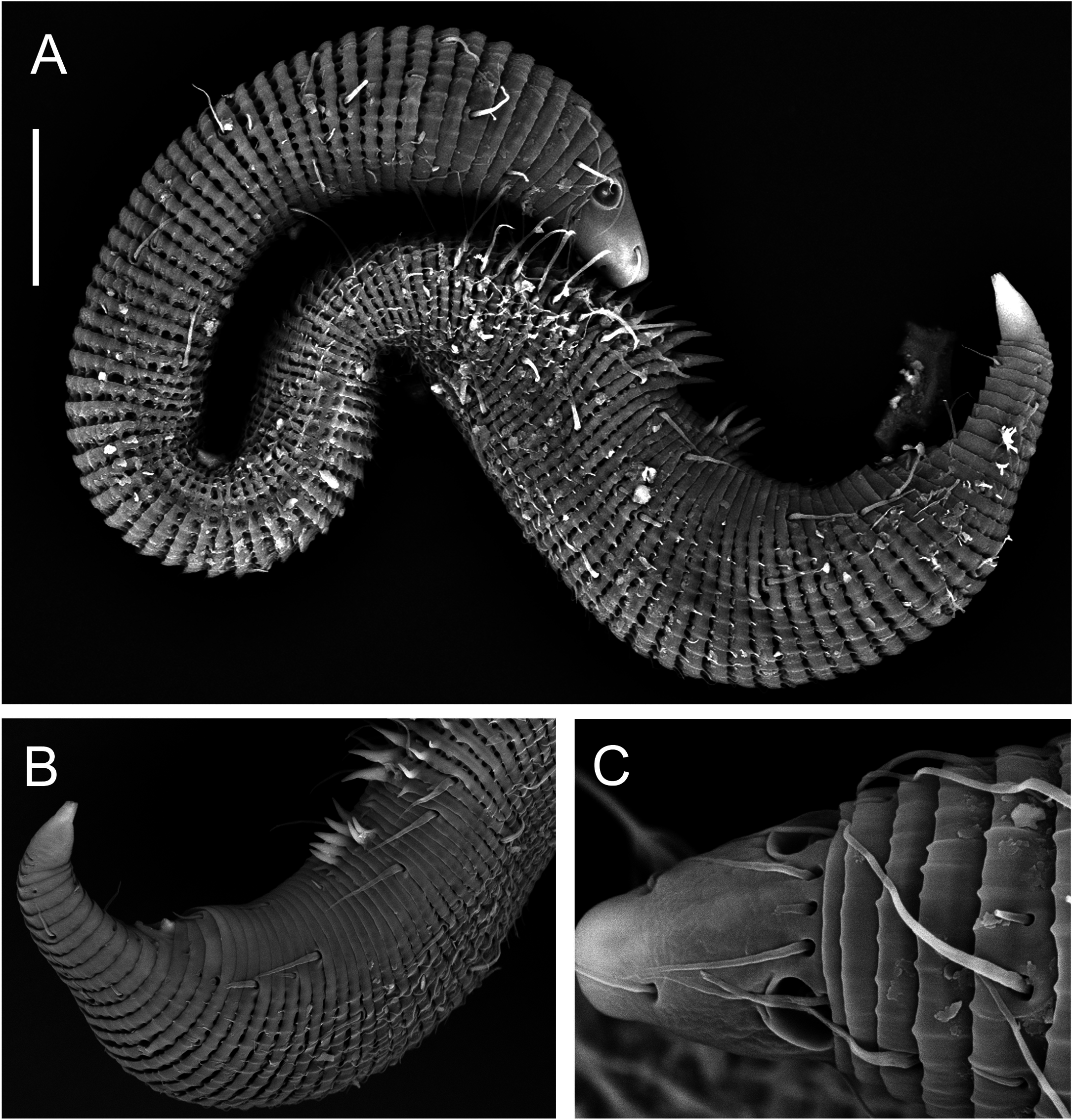
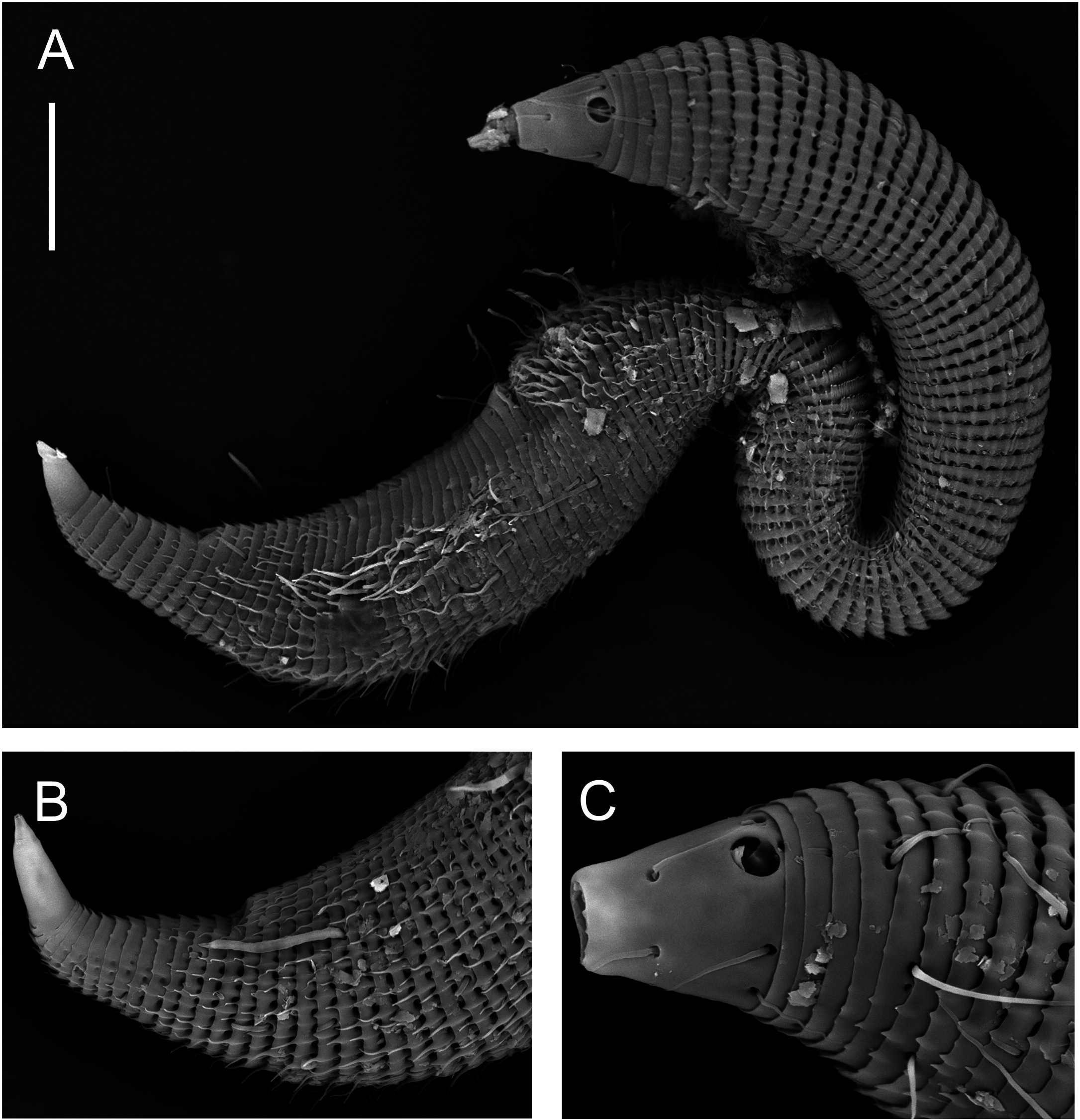
( TABLE 3, FIGS 9–13 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 )
Synonym
Epsilonema dentatum rugatum Lorenzen, 1973 .
Material observed
Five males and two females, collected on 12.i.2015, Hataitai Beach , Wellington, New Zealand (41° 18′ 22″ *At level of amphid. a, body length/maximum body diameter; abd, anal body diameter; Ant., anterior; b, body length/ pharynx length; c, body length/tail length; cbd, corresponding body diameter; diam., diameter; L, total body length; Max., maximum; Min., minimum; N, number of body rings; V, vulva distance from anterior end of body.
°S, 174° 47′ 58″ °E), intertidal coarse sand and gravel sediments ( NIWA 88387, NNCNZ 3198, 3199, 3200).
Description
Males: Body small, epsilon-shaped, with largest body diameter in posterior body region. Cuticle with 128– 132 annules overlapping with change in direction, ventrally at level of anterior dorsal curvature and dorsally at the second curvature. Cuticular ornamentation heterogeneous. Rounded vacuoles present throughout most of body, becoming progressively larger from anteriormost body ring to level of pharyngeal bulb and decreasing in size from level of cloaca towards tail tip ( Figs 12A View Figure 12 , 13B View Figure 13 ). Vacuoles largest dorsally, gradually decreasing in size ventrally, except in region of dorsal and second body curvatures where vacuoles are homogeneous; vacuoles completely absent ventrally from level of copulatory thorns to tail tip ( Fig. 12E, F View Figure 12 ). Longitudinal ridges present throughout most of body, situated between vacuoles and with similar size distribution (i.e. ridges most conspicuous where vacuoles are largest and vice versa; absent where vacuoles are absent). In body region anterior to dorsal body curvature, ridges terminate into blunt projections dorsally and into short pointed spines ventrally ( Fig. 12A, C View Figure 12 ); in region of dorsal and second body curvatures, ridges terminate into short spines, and in posterior body region, ridges terminate into longer spines gradually decreasing in length from level of cloaca towards tail tip ( Figs 11B View Figure 11 , 12F View Figure 12 ). Dense ventral rows of flattened spines present at level of, or immediately anterior to, second body curvature ( Fig. 12B View Figure 12 ). Somatic setae, 11–16 μm long, relatively sparse and irregularly arranged. Ambulatory setae with bend near base, in four longitudinal rows; two rows of four to six internal setae, and two rows of seven external setae followed posteriorly by three supporting setae. A fifth row of ambulatory setae may be present along ventral axis but could not be seen clearly with light or scanning electron microscopy.
Cuticularized rostrum, slightly longer than wide; lip region introverted. Four cephalic setae and eight subcephalic setae visible using light microscopy; an additional dorsal subcephalic seta visible using scanning electron microscopy ( Fig. 11C View Figure 11 ). Unispiral amphids situated dorsosublaterally ( Figs 11C View Figure 11 , 13C View Figure 13 ), sometimes appear circular using light microscopy. Buccal cavity small, without teeth. Pharynx with conspicuous, rounded posterior pharyngeal bulb with conspicuously cuticularized lumen. Tail short and conical, with sparse subdorsal and subventral setae ∼ 10 μm long.
Reproductive system with single anterior outstretched testis located in posterior swollen body region and situated ventrally relative to intestine. Sperm cells globular to spherical, 3–5 × 4–6 μm. Copulatory thorns beginning at level of posterior-most external ambulatory setae and comprising three longitudinal rows of six to eight thorns 3–7 μm long, becoming progressively larger posteriorly and with largest thorns located in ventral row. Seven slightly smaller precloacal thorns present, located 6–7 μm posteriorly to copulatory thorns, which, when viewed ventrally, are arranged in shape of a ‘V’ ( Fig. 12E View Figure 12 ). Spicules arcuate and with well-developed capitulum; gubernaculum short and slightly curved. Two precloacal setae present ( Fig. 11B View Figure 11 ), not observed under light microscopy.
Females: Body shape and cuticular ornamentation similar to males, but dorsal and lateral cuticular spines are conspicuously longer in posterior half of swollen posterior body region, ventral spines are present on tail and immediately anterior to cloaca, and multiple larger, ventral spines with wide bases are present in region immediately anterior to vulva ( Fig. 13A, B View Figure 13 ).
Reproductive system didelphic-amphidelphic with reflexed ovaries both located ventrally relative to intestine. Vulva located at two thirds of body length from anterior extremity. Vagina directed posteriorly, pars proximalis vaginae surrounded by constrictor muscle; vaginal glands not observed.
Remarks
The specimens described here are characterized by a greater number of body rings (128–132 vs. 124–126) and longer spicules (42–45 vs. 30–32 μm) than the Ep. dentatum rugatum specimens described by Lorenzen (1973). The size and arrangements of copulatory and precloacal thorns, and the cuticle ornamentation in both sexes, however, are identical.
Lorenzen (1973) established the distinction between Ep. dentatum and Ep. dentatum rugatum based on (1) the larger cuticular vacuoles in the latter relative to the former, and (2) the absence of transverse ridges on the body rings of Ep. dentatum . Given the large distance between the type localities of Ep. dentatum and Ep. dentatum rugatum ( Chile and New Zealand, respectively), and the importance of cuticle ornamentation for the systematics of the Epsilonematidae ( Gourbault & Decraemer, 1996) , Ep. dentatum rugatum is herein elevated to the status of species.
Based on SSU molecular sequences, Ep. rugatum Lorenzen, 1973 comb. nov. differs from Epsilonematidae sp. ( EF 591340 View Materials ) by 9% (69 in 847 bp including six gaps) and Epsilonema sp. ( FJ 182218 View Materials ) by 14% (116 in 848 bp including eight gaps) ( Fig. 14 View Figure 14 ).
PHYLOGENETIC RECONSTRUCTION
Relationships of described species
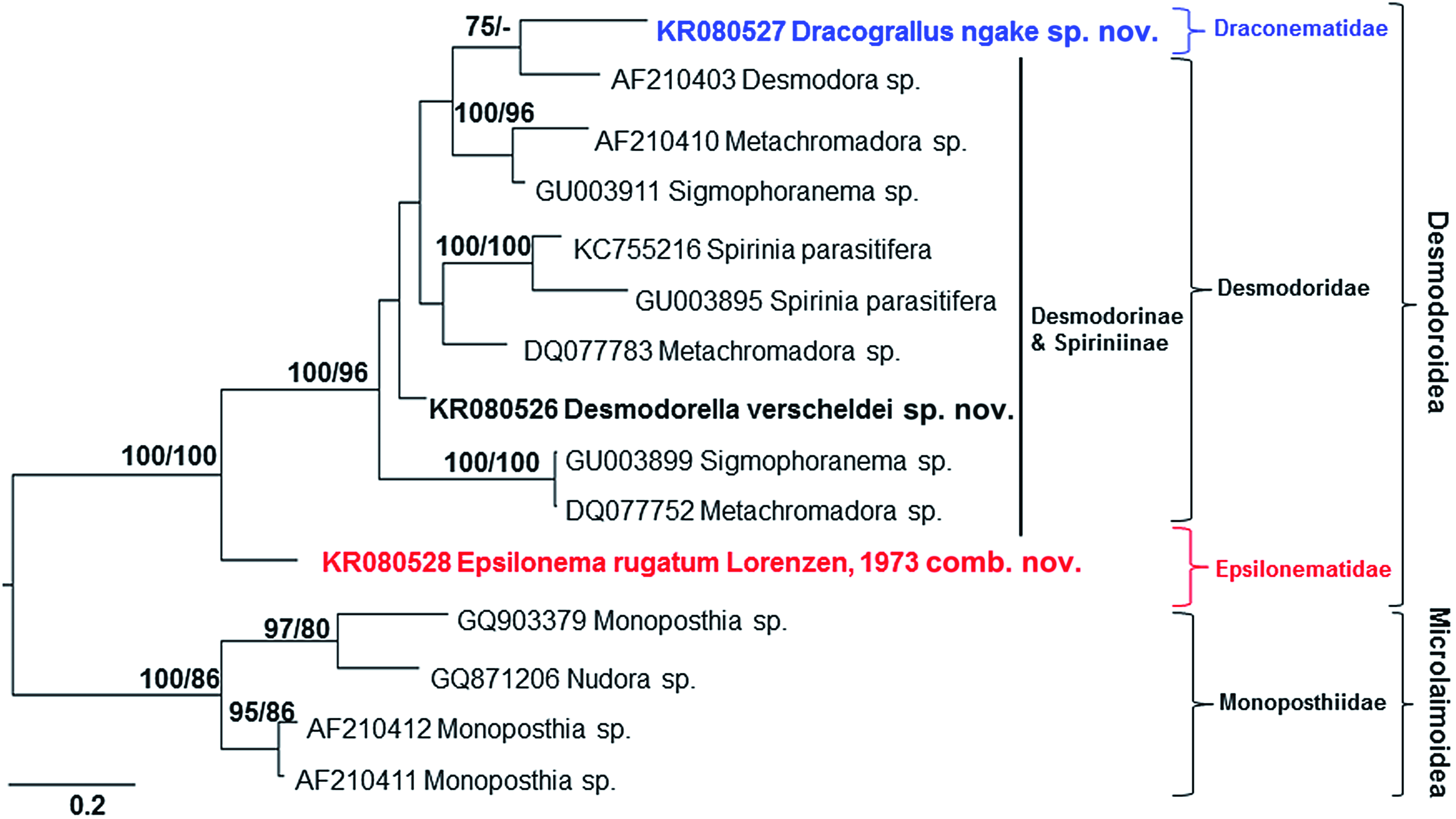
As the Bayesian and maximum likelihood trees of SSU and D2-D3 of LSU sequences were characterized by similar topologies, only the former are shown. However, bootstrap values are provided in addition to posterior probability values as a means of evaluating the validity of the results obtained using the Bayesian approach.
Desmodorella verscheldei sp. nov. was placed in a clade together with Desmodora sp. and Desmodora marci sequences in the SSU consensus tree. However, relatively low posterior probability and bootstrap values (<75%) provided only weak support for a close relationship between the two genera ( Fig. 14 View Figure 14 ). The placement of Desmodora communis well away from other Desmodora SSU sequences also suggests that the genus Desmodora is polyphyletic (assuming no misidentification, see below). The D2-D3 consensus tree did not show a close relationship between Desmodora and Desmodorella ( Fig. 15 View Figure 15 ).
Dracograllus ngakei sp. nov. and two Dracograllus sp. SSU sequences grouped together with posterior probability and bootstrap values of 100% and formed a basal clade of the Draconematidae , confirming the findings of Rho et al. (2011). The genus Prochaetosoma formed a monophyletic group, which also supports the conclusions of Rho et al. (2011). Evidence for the monophyly of this genus, however, was limited, as indicated by low posterior probability and bootstrap values. It is not possible to draw any conclusions about the relationships within the Epsilonematidae because only three SSU and one D2-D3 of LSU sequences were available.
Relationships within the Desmodoroidea based on D2-D3 and SSU phylogenetic trees
The D2-D3 and SSU consensus trees showed similar patterns ( Figs 14 View Figure 14 , 15 View Figure 15 ). Only limited inferences can be made based on the small number of available D2-D3 of LSU sequences, but our analyses suggest that: (1) Dracograllus ngakei sp. nov. is placed within a clade comprising Desmodorinae and Spiriniinae species, (2) the subfamilies Desmodorinae and Spiriniinae are paraphyletic, and (3) Ep. rugatum Lorenzen, 1973 comb. nov. forms a basal branch within the Desmodoroidea ( Fig. 15 View Figure 15 ). These patterns are discussed in more detail below in light of the more comprehensive analyses based on SSU sequences.
The SSU consensus tree shows that the Draconematidae sequences ( Dracograllus spp. , Draconema sp. Paradraconema spp. , and Prochaetosoma spp. ) formed a monophyletic group with a posterior probability of 100% and bootstrap value of 98% ( Fig. 14 View Figure 14 ). The three Epsilonematidae sequences ( Epsilonema spp. ) also formed a monophyletic group, but with lower posterior probability and bootstrap values of 97 and 55%, respectively. These results provide support for our first hypothesis, which contends that both the Draconematidae and Epsilonematidae are monophyletic, although more sequences representative of all genera (particularly for the Epsilonematidae ) will be required to provide more robust conclusions. These preliminary findings are consistent with the classification of Lorenzen (1981) based on morphological synapomorphies.
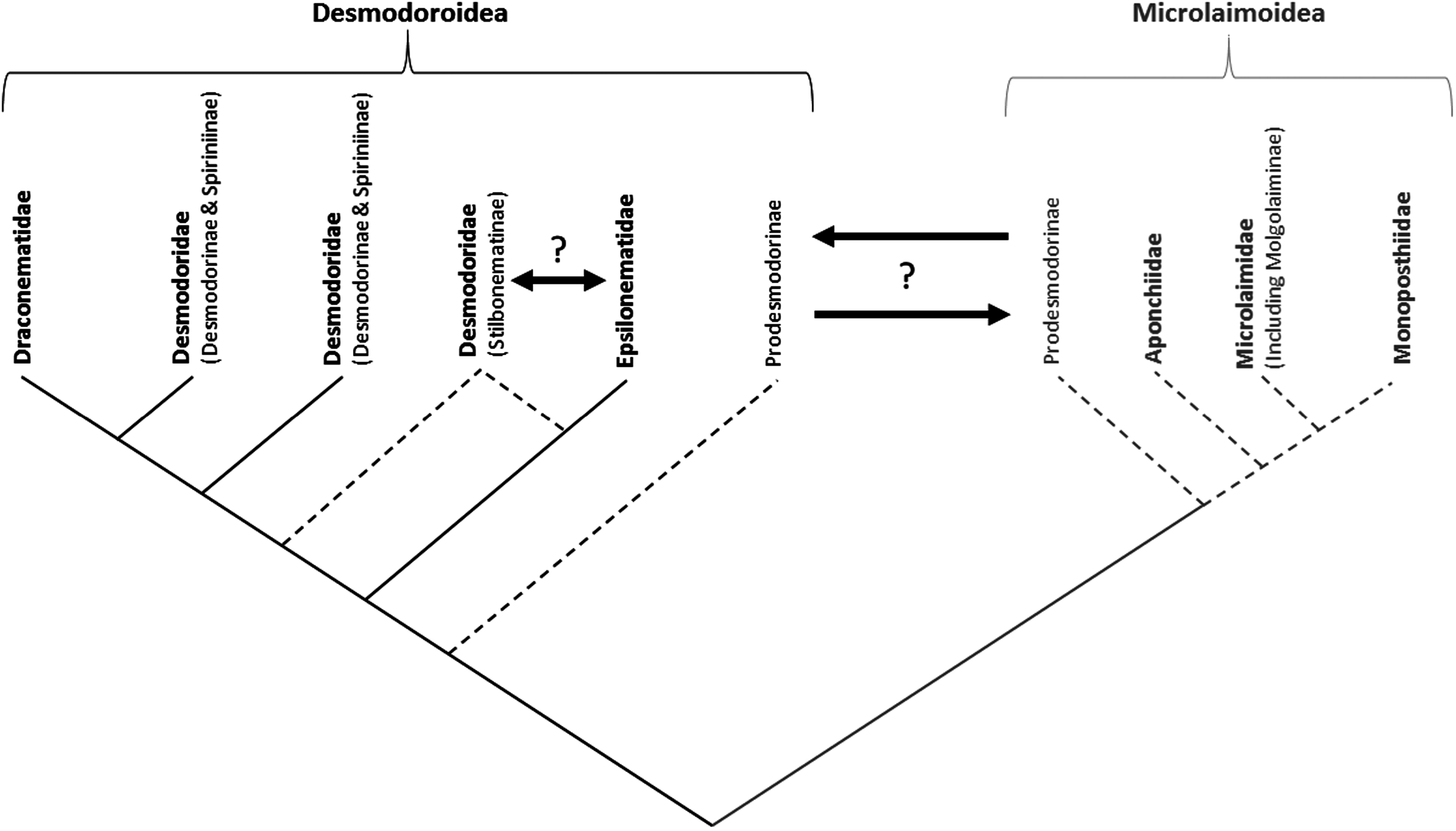
The Draconematidae and Epsilonematidae were placed in separate clades in the consensus tree. The Draconematidae formed a clade together with species of Desmodorinae and Spiriniinae , as well as a few Stilbonematinae . The placement of presumed Stilbonematinae sequences ( Robbea hypermnestra , Eubostrichus topiarius , and Robbea sp. ) together with the Desmodorinae and Spiriniinae seems unlikely, as the majority of sequences of this subfamily are on a separate branch of the tree. Two supposed sequences of Eubostrichus topiarius ( JN968219 View Materials and JN968237 View Materials ), in particular, are out of place, as another sequence of the same species ( Y16917 View Materials ) is found in a separate clade together with other Eubostrichus species. Likewise, one sequence of Robbea hypermnestra ( KJ414467 View Materials ; Ott et al., 2014) is located amongst other Stilbonematinae sequences, whereas another sequence of the same species identified by the same laboratory ( Y16921 View Materials ; Kampfer, Sturmbauer & Ott, 1998) is grouped with species of Spiriniinae . The anomalous placement of the latter species is unlikely to be a result of misidentification, and may be the result of inherent artefacts in phylogenetic tree routines ( Altaba, 2009). Therefore, our interpretation is that the family Draconematidae is a sister taxon to the Desmodorinae and Spiriniinae , with the Draconematidae forming a monophyletic crown group characterized by clear morphological synapomorphies (swollen pharyngeal body regions and mid-posterior body regions with specialized setae) whereas the Desmodorinae and Spiriniinae comprise a paraphyletic stem group without morphological synapomorphies ( Lorenzen, 1981; Fig. 16 View Figure 16 ).
The Epsilonematidae apparently formed a separate clade with species of Stilbonematinae , but the relationship between this family and subfamily remains unclear. In the consensus tree, species of Eubostrichus form a basal group to a clade comprising the Epsilonematidae and Stilbonematinae . Given the clear morphological synapomorphies characterizing the Epsilonematidae (swollen pharyngeal body regions and mid-posterior body regions with specialized setae; Lorenzen, 1981) and Stilbonematinae (glandular sensory organs and ectosymbiotic chemoautotrophic bacteria on the cuticle; Polz et al., 1992; Bauer-Nebelsick et al., 1995), we think that it is more likely that the Epsilonematidae and Stilbonematinae constitute two separate yet potentially closely related clades. The results of the present and other studies do not provide unequivocal support for the monophyly of the Stilbonematinae ( Armenteros et al., 2014a, b), but this lack of support may stem from misidentifications and artefacts of tree building. The subfamily may in fact be monophyletic, and it may be necessary to raise the Stilbonematinae to the rank of family in the future to better reflect their distinct morphology and high level of molecular divergence from subfamilies of Desmodoridae ( Figs 14 View Figure 14 , 16 View Figure 16 ).
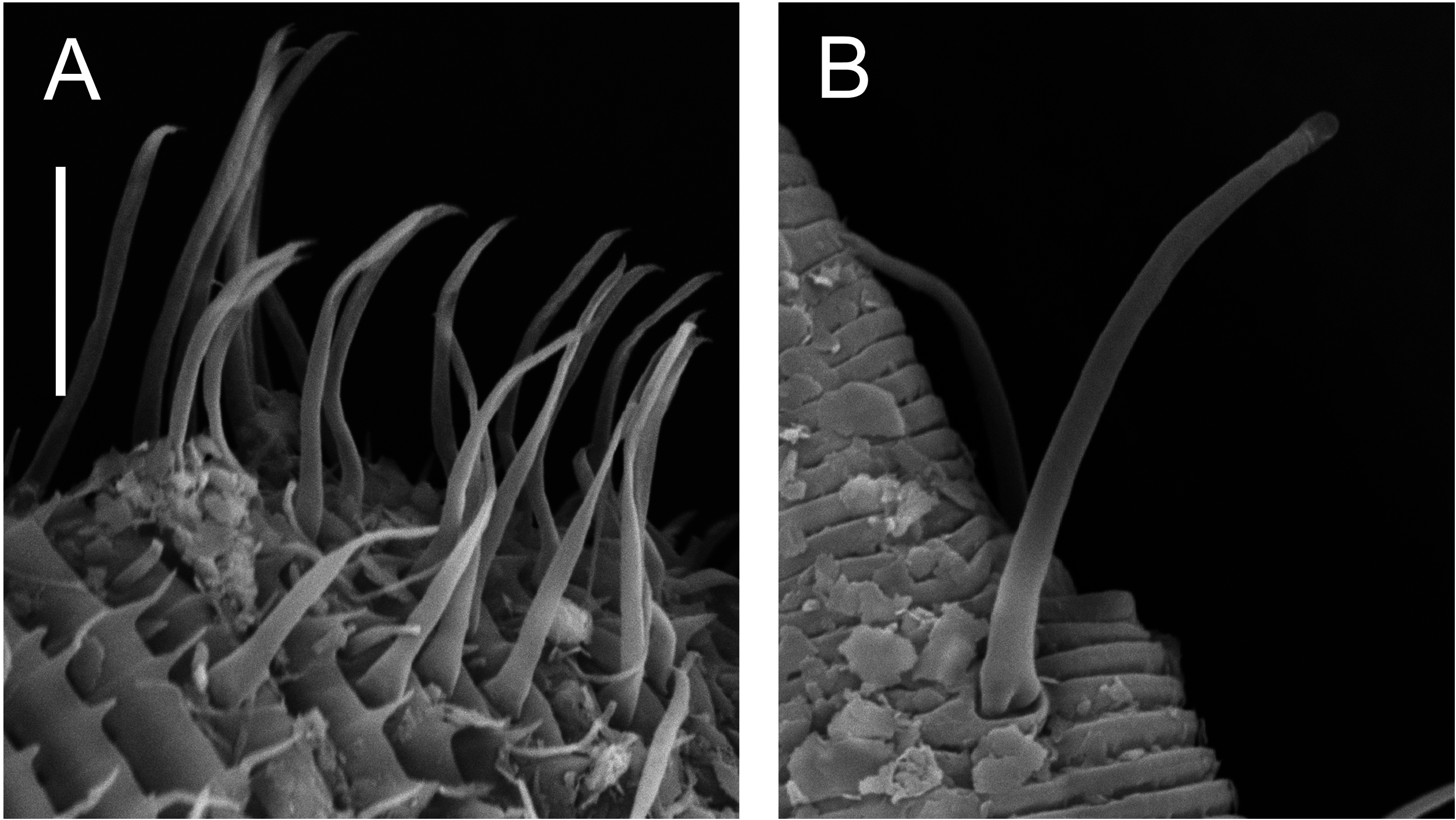
Our findings lead us to reject our second hypothesis that Draconematidae and Epsilonematidae are sister taxa. The distinctive morphological characteristics of these families (i.e. swollen pharyngeal body regions and mid-posterior body regions with specialized setae) therefore evolved independently or were present in a common ancestor and subsequently lost in Desmodoridae lineages. Although the latter is theoretically possible, recent molecular studies have demonstrated just how common convergent evolution is within the phylum Nematoda, leading van Megen et al. (2009: 927) to state that: ‘it is hard to find a morphological, ecological or biological characteristic that has not arisen at least twice during nematode evolution’. Convergent evolution would also help explain why the two families differ in two important ways. First, as noted by Verschelde & Vincx (1994), the structure of the modified setae differs markedly between the two families ( Fig. 17 View Figure 17 ). Whereas the adhesion tubes of the Draconematidae provide attachment to the substrate through epidermal gland secretions or possibly a sucker-like mechanism, the ambulatory setae of the Epsilonematidae provide traction through their claw-like distal end; the rigidity and bent shape of ambulatory setae may also provide pro- pulsion during locomotion. Second, the Draconematidae possess modified setae on the dorsal side of the anterior body region whereas none are found in the Epsilonematidae . The morphological similarities between the two families would therefore appear to reflect similar, but distinct and independently evolved adaptations to their unusual mode of locomotion, which consists of a caterpillar-like walking motion as opposed to serpentine coiling in all other nematodes.
Our analyses clearly show that the family Desmodoridae is paraphyletic, which is contrary to the conclusion of Armenteros et al. (2014a), but consistent with the more comprehensive analyses of Meldal et al. (2007) and van Megen et al. (2009). As noted by the latter authors, the highly divergent position of Prodesmodora puts into question whether the Prodesmodorinae should indeed reside in the family Desmodoridae . Based on our results, and those of van Megen et al. (2009), we propose that the Prodesmodorinae be removed from the Desmodoridae . Defining the status and relationships of this group, however, will require a wider molecular investigation of lineages related to the Desmodoroidea ; relationships with the Desmodoroidea or Microlaimoidea both appear possible at present. We note that the Prodesmodorinae are found in terrestrial or freshwater environments exclusively, suggesting that the genetic divergence of this group amongst predominantly marine lineages may be the result of adaptation to a different environment ( van Megen et al., 2009).
The placement of Molgolaimus demani with species of Microlaimidae indicates that the Molgolaiminae should be classified with the Microlaimoidea instead of the Desmodoroidea , and that the latter superfamily is therefore paraphyletic ( Figs 14 View Figure 14 , 16 View Figure 16 ; Meldal et al., 2007). Molgolaimus Ditlevsen, 1921 , was classified with the Microlaimidae (see Gerlach & Riemann, 1973 /1974), based on similarities to the genus Microlaimus De Man, 1880 , in head and amphideal fovea shape, arrangement of head sensillae, and buccal cavity structure. Jensen (1978) later erected the family Molgolaimidae Jensen, 1978 , distinguished from the Microlaimidae mainly based on the presence of a single anterior testis in males and two reflexed ovaries in females. Most recently, Lorenzen (1981) placed Molgolaimus in its own single-genus subfamily within the Desmodoridae and established the monophyly of the Desmodoroidea based on the presence of only one (anterior) testis in males. It appears that in the case of the Desmodoroidea and Microlaimoidea the structure of the gonads is not a reliable trait to use for classification. We note however that molecular sequences are only available for one Molgolaimus species , and that additional data for the genus are needed to verify its relationships.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Epsilonema rugatum
| Leduc, Daniel & Zhao, Zeng 2016 |
Robbea hypermnestra
| Ott, Gruber-Vodicka, Leisch & Zimmermann 2014 |
Robbea hypermnestra
| Ott, Gruber-Vodicka, Leisch & Zimmermann 2014 |
Eubostrichus topiarius
| Berger, Urbancik & Ott 1996 |
Eubostrichus topiarius
| Berger, Urbancik & Ott 1996 |
Epsilonema dentatum rugatum
| Lorenzen 1973 |
Epsilonema rugatum
| Lorenzen 1973 |
Spiriniinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Spiriniinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Spiriniinae
| Chitwood 1936 |
Spiriniinae
| Chitwood 1936 |
Spiriniinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Stilbonematinae
| Chitwood 1936 |
Desmodorinae
| Filipjev 1922 |
Desmodorinae
| Filipjev 1922 |
Desmodorinae
| Filipjev 1922 |
Desmodorinae
| Filipjev 1922 |
Desmodoridae
| Filipjev 1922 |
Desmodoridae
| Filipjev 1922 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |
Draconematidae
| Filipjev 1918 |