
Brachyorrhos wallacei, Murphy & Mumpuni & Lang & Gower & Sanders, 2012
|
publication ID |
https://doi.org/10.5281/zenodo.5350190 |
|
persistent identifier |
https://treatment.plazi.org/id/E37587A2-7E7B-FFD9-FF21-E977FD355D7B |
|
treatment provided by |
Tatiana |
|
scientific name |
Brachyorrhos wallacei |
| status |
sp. nov. |
Brachyorrhos wallacei new species
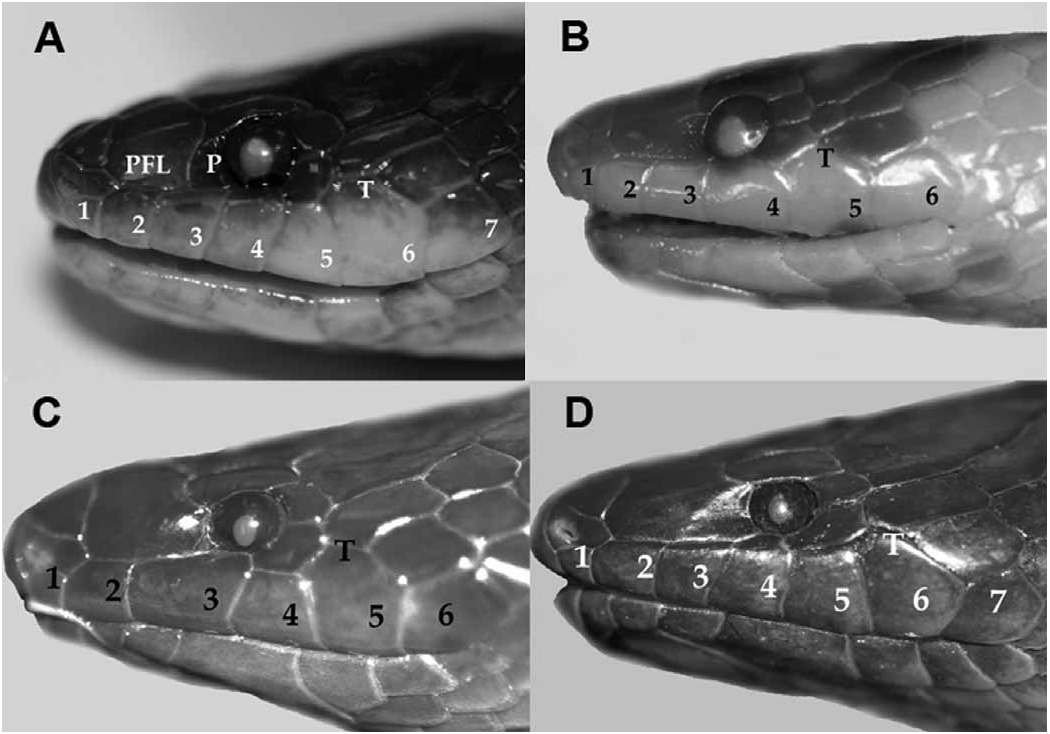
( Fig. 4d View Fig )
Brachyorrhos albus Boulenger, 1893: 305
Brachyorrhos albus Setiadi & Hamidy, 2006 (two un-numbered photographs, no pagination)
Material Examined. — Holotype. MZB 3463 View Materials an adult female with a total length of 722 mm from Jailolo, Halmahera. This new species is based on 10 adults from the island of Halmahera ( Jailolo – MZB 3464 View Materials , MZB 3462 View Materials ; Jailolo , Kampung PasirPatih – USNM 237145–150 About USNM ; Sidangoli – MZB 2111 View Materials ; Weda Bay Mines – MZB 4487 View Materials .)
Diagnosis. — A Brachyorrhos with no preocular, usually seven (rarely six) upper labials, the tallest of which is six (rarely five); ventrals 177–182, subcaudals 38–42. This snake is very similar to B. raffrayi which has upper labial three in the orbit and the third is the tallest. Specimens of wallacei usually have the fourth upper labial (rarely 3 + 4) in the orbit, and the sixth is the tallest. Our mitochondrial data show that B. raffrayi and B. wallacei are separated by a deep molecular divergence at this locus.
Description of Holotype. — An adult female 660 mm SVL, 82 mm tail. Rostral visible from above, internasals paired, prefrontal fused with loreal and preocular; nasal scales bilobed with posterior lobe dorsal to the first labial; frontal longer than seam of parietals; parietals extend laterally to make contact with upper postocular; two postoculars; one primary temporal scale contacts the lower postocular, a secondary temporal on each side contacts upper labial six; seven upper labials, 2–4 make contact with prefrontal, the fourth enters the orbit, the fifth is the tallest; seven lower labials, the first four make contact with the chin shields; dorsal scales rows are 19 at mid-body; ventrals 180; subcaudals 42. Dorsum grey, anterior body has scale rows one and two with red-brown pigment, replaced by grey posteriorly, ventral surface a uniform red brown.
Paratypes. — The nasals are divided or semi-divided with the nostril in the anterior portion of the scale. Primary temporal scale(s) can be one or two and plate-like. Upper labials number six (or seven), 2-3 (or 2-4) make contact with the fused loreal-prefrontal shield; when the third upper labial enters the orbit the fifth upper labial is the tallest; when the fourth upper labial enters the orbit the sixth is the tallest. One specimen (MZB 3464) has the postoculars fused on the left side. Lower labials seven, the first pair of lower labials make contact on the midline of the chin posterior to the mental. The first four contact the only pair of chin shields. Dorsal scales on the body are smooth and in 19 rows, the number of rows at the 10 th ventral may be 19 or 20. Males have 169–182 (N = 7, = 177.14, SD = 4.25) ventrals and 33–42 (N = 7, = 38.5, SD = 3.35) subcaudals. Females have 170–174 (N = 3, = 171.3, SD = 1.88) ventrals and 26–31 (N = 3, = 28.6, SD = 2.04) subcaudals.
In alcohol, grey-black above with some yellow under the chin; Setiadi and Hamidy (2006) contains two photographs of a live specimen of this species, showing a uniform dark brown-purple snake with a white venter. The specimen in their photo has 7 upper labials, with 3 rd upper labial in the orbit.
Size. — Males have SVLs of 453–660mm (N = 7, = 561.8, SD = 67.7); tails that are 56–82 mm (n = 7, = 71, SD = 10.03), and t/svl ratios 12.0–13.4 (N = 7, = 12.67, SD = 0.47). Females have SVLs of 394–462 mm (N = 3, = 417.0, SD = 31.8) and tails that are 29–51 mm (= 38.0, SD = 9.41); tail/SVL ratios 7–11% (N = 7, = 9%, SD = 0.015).
Etymology. — Named in honour of Alfred Russell Wallace for his contributions to the natural history of Halmahera.
Unresolved Brachyorrhos . — (1) The Aru Islands specimen clearly belongs with this genus despite the speculation that it is jobiensis (Iskandar & Colijn, 2001) . It clusters with the Buru island gastrotaenius in our PCA and cluster analysis. Its locality data may be in error. (2) The island of Buru appears to have a second species with distinct labial architecture and a long tail. However, this specimen is poorly preserved and more sampling is needed to inform its status. (3) Seram also supports a second species of Brachyorrhos that is exceptionally small (smaller than late term embryos of B. albus ) and sexually mature with a distinct loreal scale, preocular, and low ventral count. These specimens will be discussed elsewhere pending further investigation. (4) Morotai also supports a population of Brachyorrhos (USNM 159973) that is superficially similar to B. raffrayi , with a slightly different labial architecture, and it may require molecular analysis to confirm its relationships. (5)
Brachyorrhos has also been reported from: Banda, Batjan,
Haruku, Nusalaut, and Saparua; all of these islands seem likely to support Brachyorrhos populations, but we have not located specimens to confirm their presence or identities.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Brachyorrhos wallacei
| Murphy, John C., Mumpuni, Lang, Ruud de, Gower, David J. & Sanders, Kate L. 2012 |
Brachyorrhos albus
| Boulenger, G 1893: 305 |