
Gilmoremys, Joyce & Lyson, 2011
|
publication ID |
https://doi.org/10.1080/14772019.2023.2217505 |
|
publication LSID |
lsid:zoobank.org:pub:E8E5C4DE-E52A-43FB-B9C1-CE3ED2ED66C1 |
|
DOI |
https://doi.org/10.5281/zenodo.10980327 |
|
persistent identifier |
https://treatment.plazi.org/id/E459E443-FF81-5842-FCEF-A104FACFED88 |
|
treatment provided by |
Felipe |
|
scientific name |
Gilmoremys |
| status |
|
Gilmoremys from the Cretaceous of North America
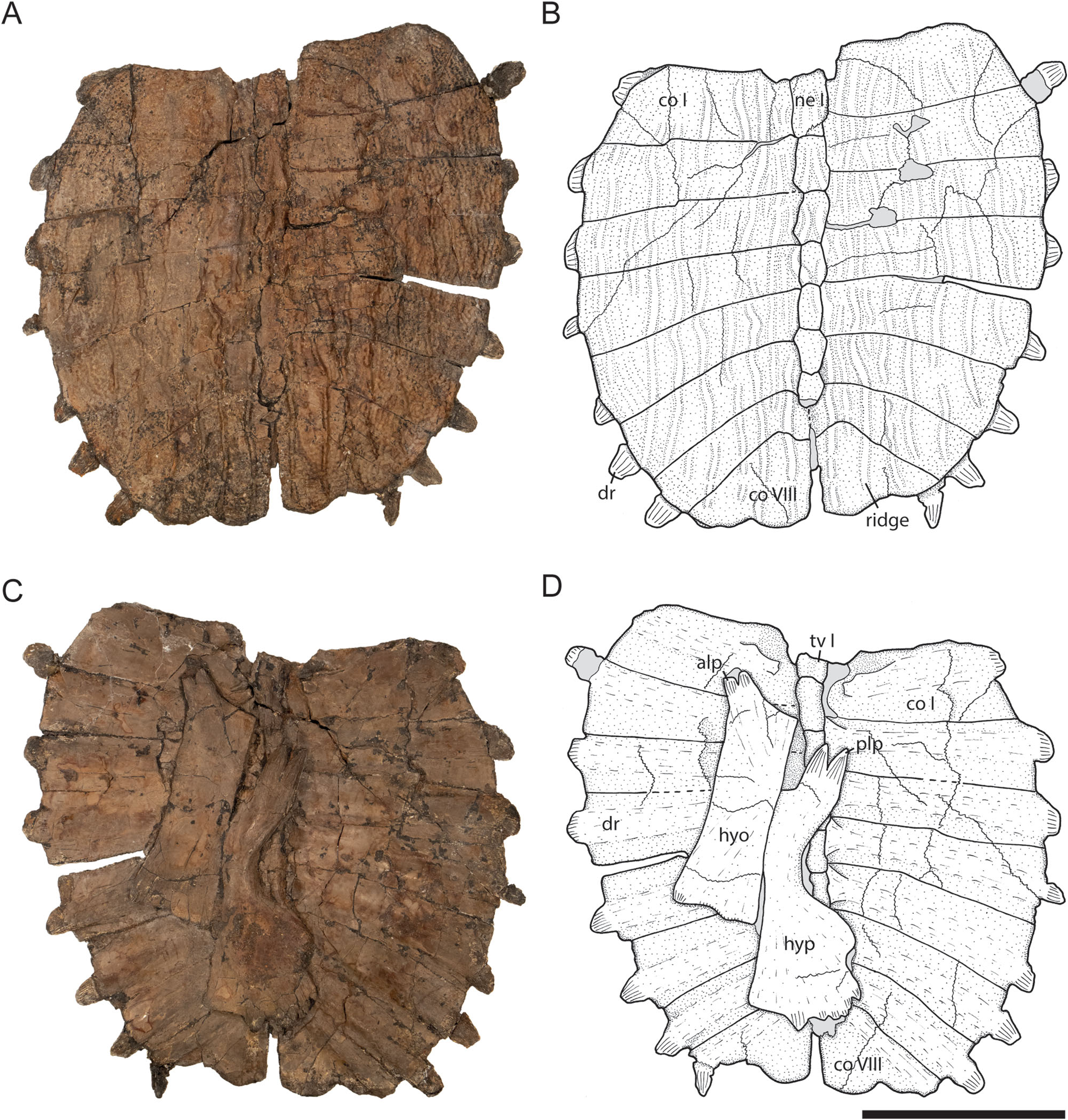
Besides S. impressa , the closest resemblance to S. baba among Pan-Trionychidae is shown by some members of Plastomenidae from the Late Cretaceous of North America. The carapace and plastron of Gilmoremys lancensis ( Gilmore, 1916) and especially of Gilmoremys gettyspherensis Joyce, Lyson and Sertich, 2018 are very similar in overall morphology to both species of Striatochelys .
Carapace. Gilmoremys lancensis has an overall carapace length of 34 cm and thus is larger than both S. baba and S. impressa (carapace lengths of 27 cm and 14 cm, respectively) as well as its sister taxon G. gettyspherensis (carapace length of 25 cm). The overall carapace shape is similar in G. lancensis and G. gettyspherensis , as well as in S. baba and S. impressa , all having an oval outline and a distinctive sculpturing on the dorsal surface of the carapace. G. lancensis differs from both G. gettyspherensis and the two species of Striatochelys in having sculpturing that consists of grooves ( Joyce & Lyson, 2011, figs 8, 10). In G. lancensis , S. baba , and S. impressa , on the other hand, the sculpturing consists of well-developed longitudinal ridges, that reach from costal I anteriorly to costal VIII posteriorly (for S. baba , see Figs 2 View Figure 2 , 3 View Figure 3 ; for S. impressa see Yeh [1963, pl. XIX, fig. 3, and pl. XX, fig. 1]; for G. gettyspherensis see Joyce et al. [2018, fig. 2]).
The nuchal of G. lancensis is as wide as it is in Striatochelys . It differs from the latter, however, in the absence of a nuchal emargination ( Joyce & Lyson, 2011, figs 8, 10). Further differences occur in the number of neurals. Striatochelys baba and G. lancensis both have a total of seven neurals, whereas there are eight neurals present in S. impressa and G. gettyspherensis . Another difference lies in the presence of a preneural and a small first neural in the two species of Gilmoremys ( Joyce & Lyson, 2011, figs 8, 10; Joyce et al., 2018, fig. 2). In S. baba , the preneural is absent ( Figs 2 View Figure 2 , 3 View Figure 3 ); in S. impressa , the region is not preserved. In Gilmoremys , costal II is laterally enlarged and projects anteriorly, reducing the lateral margin of costal I ( Joyce & Lyson, 2011, figs 8, 10; Joyce et al., 2018, fig. 2). In Striatochelys , by contrast, the distal part of costal II is not enlarged and the distal margin of costal I and II are similarly-sized ( Figs 2 View Figure 2 , 3 View Figure 3 ; Ye, 1994, fig. 70). Costal VIII is, in all four species, triangular and enlarged. It is largest in S. baba , in which it is anteroposteriorly slightly longer than costal VI and VII together, whereas it is barely the size of costal VI and VII together in the other species.
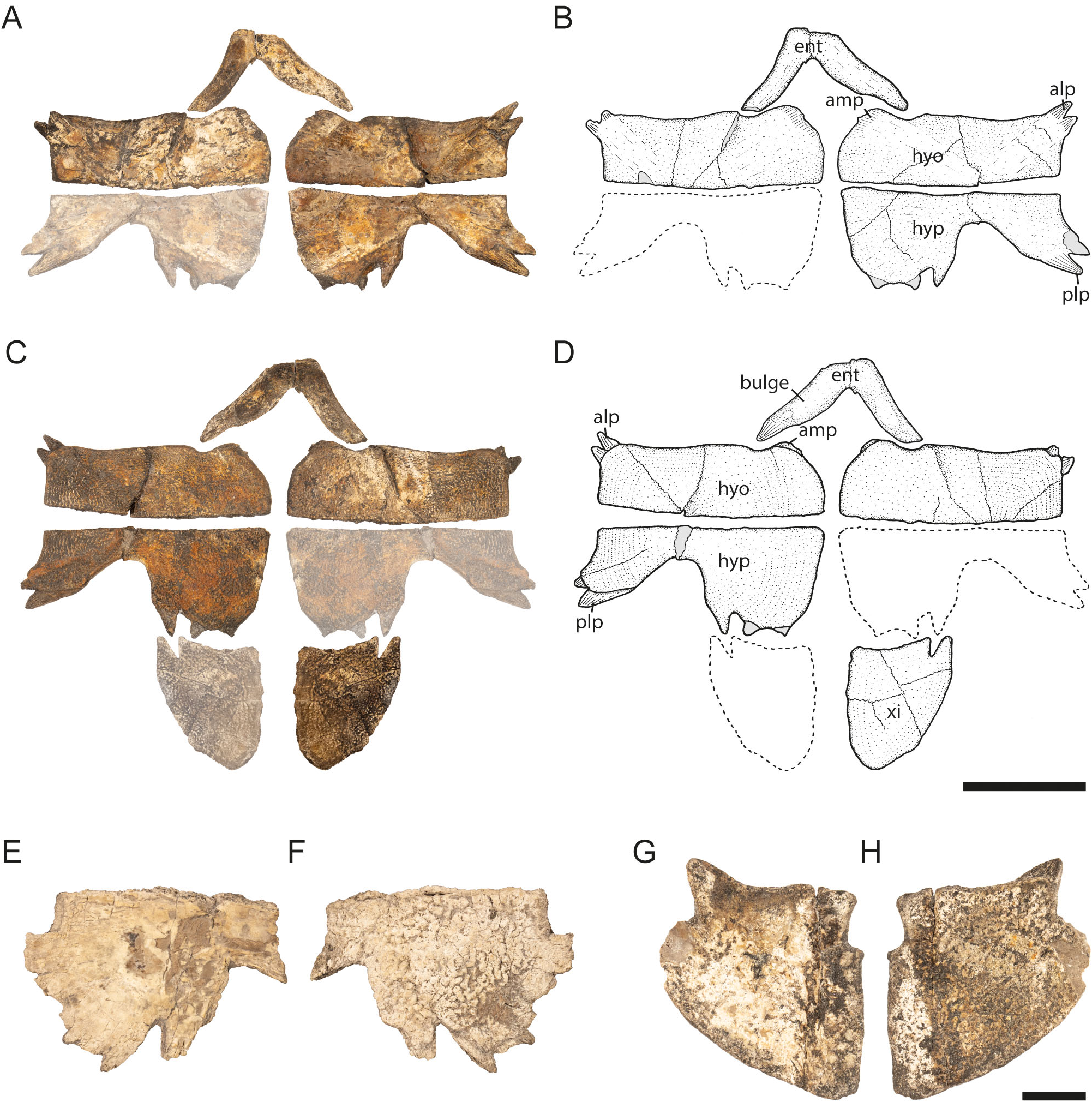
Plastron. In contrast to the carapace, which shows many (small) differences despite its overall resemblance, the plastron is extremely similar in Striatochelys and Gilmoremys . The hyoplastron of Gilmoremys has a well-developed anterior notch for the entoplastron ( Joyce & Lyson, 2011, figs 9, 11; Joyce et al., 2018, fig. 3), a feature that is also present in one individual of S. baba ( Fig. 5 View Figure 5 ), but absent in two others ( Figs 3 View Figure 3 , 4G, H View Figure 4 ). The posterolateral processes of the hypoplastron of Gilmoremys are slightly larger than in S. baba , while the fan-like posteromedial processes present in Striatochelys are only weakly developed or absent in Gilmoremys ( Joyce & Lyson, 2011, figs 9, 11; Joyce et al., 2018, fig. 3). The xiphiplastron in all three species has the same triangular morphology but is anteroposteriorly slightly longer in Gilmoremys ( Joyce & Lyson, 2011, figs 9, 11; Joyce et al., 2018, fig. 3) than in S. baba ( Figs 5C, D, G, H View Figure 5 ).
Extant taxa from Southeast Asia
Southeast Asia has the highest diversity of extant trionychids. According to molecular data, most Asian taxa belong to a single monophyletic group, including five genera, i.e. Pelodiscus , Palea , Dogania , Amyda and Nilssonia , which likely originated between the Eocene and Miocene ( Engstrom et al., 2004; Le et al., 2014; Pereira et al., 2017; Thomson et al., 2021). Based on the middle to late Eocene age and Southeast Asian distribution of Striatochelys baba , a comparison with selected members of this extant group is provided below.
Carapace. With a bony disc length (BDL) of 27 cm (GPIT-PV-122875), S. baba is far larger than the smallest Pelodiscus species, i.e. Pelodiscus huangshanensis and Pelodiscus shipian , which barely reach 10 cm BDL, but it is much smaller than the large Nilssonia and Amyda species (e.g. Nilssonia leithi [ Gray, 1872], Nilssonia gangetica [ Cuvier, 1825], Amyda cartilaginea [ Boddaert, 1770]), with a BDL between 38 and 60 cm. In contrast to this, Dogania subplana ( Geoffroy Saint-Hilaire, 1809) and Palea steindachneri ( Siebenrock, 1906) have a roughly similar size, with 21.7 and 30 cm BDL, respectively (S. Gong et al., 2022; Y. A. Gong et al., 2021; Pritchard, 2001). The overall carapace morphology in different taxa of Pan-Trionychinae is very similar, but one notable difference is the presence or absence of multiple anteroposteriorly extending ridges across the carapace. Such ridges are prominent in S. baba (see above). In extant eastern Asian taxa, similar but weaker ridges are often found in early ontogenetic stages (e.g. in Pelodiscus sinensis ; IVPP 525, USNM 539334). If such ridges are also present later in ontogeny (e.g. in A. cartilaginea ; FMNH 11088, USNM 22522, 222521), they are much more weakly developed in comparison to S. baba and never appear as double rows on the neurals.
In S. baba , the nuchal is about four times wider than it is long. Among extant Eastern Asian taxa, this is only the case in the genus Pelodiscus View in CoL (e.g. P. sinensis View in CoL and Pelodiscus jiangxiensis Hou et al., 2021 ) and in D. suplana , whereas in other taxa the nuchal is only two to three times wider than long. The absence of a preneural, as in S. baba ( Figs 2 View Figure 2 , 3 View Figure 3 ), is common in extant Eastern Asian taxa. Notable exceptions to this are two species of the genus Nilssonia View in CoL , i.e. Nilssonia hurum ( Gray, 1831) View in CoL and N. gangetica View in CoL , where a separated preneural is formed. Striatochelys baba has seven neurals ( Figs 2 View Figure 2 , 3 View Figure 3 ). In extant taxa, the number of neurals is often variable within a given genus or species, ranging from seven to eight. However, some taxa of the eastern Asian group ( D. subplana View in CoL , A. cartilaginea View in CoL , Nilssonia formosa View in CoL [ Gray, 1869] and N. hurum View in CoL ) invariably have eight neurals. The point of reversal for the neural orientation is less variable in extant eastern Asian taxa and is usually at the fifth or sixth neural. Only N. gangetica View in CoL and N. hurum View in CoL show the same intraspecific variability as S. baba ( Figs 2 View Figure 2 , 3 View Figure 3 , 4G View Figure 4 ), in which the reversal occurs at either the fifth or the sixth neural. In S. baba , costals I–VII are relatively uniformly shaped ( Figs 2 View Figure 2 , 3 View Figure 3 ). This is also the case for most extant eastern Asian taxa, but in D. subplana View in CoL ( FMNH 224111, USNM 222523, UCMVZ 95937) and N. leithii View in CoL ( FMNH 224231), costal II is distally expanded, similar to the morphology of D. tethyensis and ‘ T. ’ ninae (see above). Remarkably, S. baba has an enlarged triangular costal VIII, which is longer anteroposteriorly than costal VI and VII combined ( Figs 2 View Figure 2 , 3 View Figure 3 ). In Nilssonia spp. (NHMUK 86.8.26.2, FMNH 223231) and A. cartilaginea View in CoL ( FMNH 11088, USNM 22522), costal VIII is also triangular and thus shaped similarly, but somewhat smaller than in S. baba , while in D. subplana View in CoL , costal VIII has the same elongated morphology as costals I–VII.
Plastron. As mentioned above, the plastron of S. baba is relatively well developed and robust. In some extant east Asian species, i.e. A. cartilaginea and D. subplana , the plastron is much thinner and the hyo- and hypoplastra are much further apart from their counterparts. In other species, i.e. Nilssonia spp. and P. sinensis , however, the overall plastron morphology is very similar to that of S. baba .
The entoplastron of S. baba is relatively slender, it possesses a callosity in the shape of a bulge and its branches project into a notch of the hyoplastron ( Fig. 5 View Figure 5 ). In P. sinensis ( IVPP 556, USNM 539335), the entoplastron is much larger and its branches are slenderer. Additionally, callosities are much less developed and the branches do not project into a hyoplastral notch. In A. cartilaginea ( FMNH 11088, USNM 22522) and D. subplana ( FMNH 224111, USNM 222523), the general shape of the entoplastron resembles the condition in S. baba , but callosities are much less developed and the branches do not extend into a hyoplastral notch. In N. gangetica , the entpoplastron is either similarly shaped ( FMNH 260430, USNM 293693) or slightly slenderer ( NHMUK 86.8.26.1) than in S. baba , the callosities are much more weakly developed and the branches do not project into a hyoplastral notch. In N. hurum ( NHMUK 86.8.26.2), the entoplastron has a shape similar to that in S. baba and also shows a callosity in the shape of a bulge. However, the bulge is much more weakly developed than that of S. baba . Furthermore, the branches do not project into a hyoplastral notch. In N. leithi ( FMNH 224231), the shape is again similar to S. baba , but callosities are less developed. In contrast to the extant taxa mentioned above, N. leithi shows well-defined hyoplastral notches for the branches of the entoplastron.
The hyoplastron of S. baba has a moderately short pair of anterolateral processes and at least two short processes anteromedially, which appear to be even shorter than the lateral ones, but their surface is abraded ( Figs 3 View Figure 3 , 5 View Figure 5 ). The morphology of the hyoplastron differs from that found in A. cartilaginea ( FMNH 11088, USNM 22522) and D. subplana ( NMNH 222523, UCMVZ 95937), in which the anterolateral processes are much longer and the anteromedial processes are both much longer and more numerous. In P. sinensis ( IVPP 556, USNM 68476), the hyoplastron is much more similar to that of S. baba . The anterolateral and anteromedial processes are still longer than in S. baba , but much shorter than in the aforementioned two taxa. In N. gangetica (NHMUK 86.8.26.1, USNM 293693), N. hurum ( NHMUK 86.8.26.2) and N. leithi ( FMNH 224231), the processes are more or less identical to those of S. baba . Possible small differences in the anteromedial processes can easily be explained by the abraded processes of S. baba .
A similar picture emerges in the hypoplastron. The processes in S. baba are short and similar in length to those in the hyoplastron. As in the hyoplastron, A. cartilaginea ( FMNH 11088, USNM 22522) and D. subplana ( USNM 222523, UCMVZ 95937) show a different morphology with much longer processes, whereas the morphology in N. gangetica (NHMUK 86.8.26.1, NHMUK 293693), N. hurum ( NHMUK 86.8.26.2) and N. leithi ( FMNH 224231) is almost identical to that found in S. baba . Pelodiscus sinensis ( IVPP 556, USNM 68476) is again intermediate in this respect: its processes are shorter than in A. cartilaginea and D. subplana , but still longer than in Nilssonia spp. and S. baba .
The xiphiplastron of S. baba is relatively large, triangular and has short abraded processes ( Figs 2 View Figure 2 , 5 View Figure 5 ). As with the hyo- and hypoplastron the closest similarities are present within Nilssonia spp. and S. baba . In N. gangetica (NHMUK 86.8.26.1, USNM 293693) and N. hurum ( NHMUK 86.8.26.2) the xiphiplastron looks almost identical to that found in S. baba . Small differences occur in the medial processes, which are slightly more pronounced and are not covered with callosities in S. baba . In N. leithi ( FMNH 224231), the xiphiplastron is more massive than in S. baba and anteroposteriorly almost as long as the hyo- and hypoplastron combined, whereas in S. baba the xiphiplastron reaches only the anteroposterior length of the hypoplastron. In P. sinensis ( IVPP 556, USNM 68476), the xiphiplastron is less triangular and more elongated. In addition, there is no direct medial contact between the left and right counterparts as is the case for Nilssonia spp. and probably also for S. baba . In A. cartilaginea ( FMNH 11088, USNM 22522) and D. subplana ( USNM 222523, UCMVZ 95937), the xiphiplastron is much slenderer and anterolaterally wider than in S. baba .
Discussion of the comparisons
Comparisons with the Palaeogene taxa from Asia underline a general issue concerning Pan-Trionychidae: despite many occurrences, the material in question often consists only of fragmentary shell remains, which are rarely diagnostic at the species level (for discussion of the issue, see also Georgalis & Joyce [2017]). Only eight Palaeogene Asian species were considered valid by Georgalis and Joyce (2017), but their preservation differs greatly. Whereas there are multiple complete individuals preserved for ‘ Trionyx ’ gregarius, including at least one juvenile, only a single almost complete carapace has been described for Drazinderetes tethyensis , ‘ Trionyx ’ johnsoni, ‘ Trionyx ’ ninae (including some additional fragmentary material) and Striatochelys impressa . The only specimen of Kuhnemys palaeocenica is a nearly complete shell of a juvenile individual. On the other hand, ‘ Trionyx ’ linchuensis only preserves an anterior carapace fragment, and for ‘ Trionyx ’ minusculus only a hyo- and hypoplastron are known. Pan-Trionychidae are, however, known for their high intraspecific variation ( Meylan, 1987), making comparisons based on single individuals especially difficult.
Aside from the aforementioned Asian Pan-Trionychidae , we also provided a comparison with Plastomenidae due to it being the only group with prominent anteroposteriorly extending ridges as adults and a very similar plastron morphology; this is especially true for the Late Cretaceous genus Gilmoremys . However, plastomenids are so far known only from the Cretaceous and Palaeogene of North America ( Joyce & Lyson, 2011; Joyce et al., 2018).
Overall, three features of the carapace and plastron are potentially important for a discussion of the close relationship between S. baba and S. impressa , as well as to untangle the potential affinity of Striatochelys within either Pan-Trionychinae or Plastomenidae .
1. Multiple anteroposteriorly projecting ridges on the carapace are rare in Pan-Trionychidae. They occur in some individuals of extant species (e.g. Apalone ferox, AMNH 57384, AMNH 65622; Trionyx triunguis ( Forskål, 1775) , AMNH 50723, AMNH 50724; Amyda cartilaginea, FMNH 11088, USNM 22522, 222521; Pelodiscus sinensis, IVPP 525, USNM 539334), but are much weaker, usually only present in juvenile/subadult individuals and only rarely present in adult specimens. In fossil taxa, however, they also sometimes occur in the shape of prominent ridges in adult specimens. Besides Striatochelys , they also appear in Plastomenus , e.g. Plastomenus vegetus ( Gilmore, 1919) , in Gilmoremys gettyspherensis and (in the form of a finer striation present only posteriorly) in ‘ T. ’ gregarius. All species that have prominent carapacial ridges as adults thus belong to Plastomenidae . Although such ridges are absent in adult pantrionychines, the appearance of such ridges in juveniles of several (not particularly closely related) trionychine species, indicates that some species might have retained this morphology in later ontogenetic stages as well.
2. The difference in the number of neurals between S. baba and S. impressa is of low significance for separating these taxa. It is known that the number of neurals varies between individuals of a single species in Pan-Trionychidae ( Meylan, 1987). Additionally, the number of individuals showing a complete row of neurals is too low in both taxa (three individuals in S. baba , one individual in S. impressa ) to establish a clear difference in this regard. The absence of a preneural (first neural fused to the second neural in the character-taxon matrix of Meylan, 1987) is exclusively known for Pan-Trionychinae, with a reversal in Nilssonia gangetica and Nilssonia hurum and basal trionychids. The absence of a preneural in S. baba hence strongly supports a position for Striatochelys inside Pan-Trionychinae. Costals I and II are similarly shaped in S. baba and S. impressa , as in other Asian Pan-Trionychidae, except for D. tethyensis and ‘ T. ’ ninae. This stands in marked contrast to the condition present in Gilmoremys , in which costal II is strongly bowed anterolaterally, further pointing away from a close relationship between Striatochelys and Gilmoremys .
3. The plastron of S. baba is remarkably similar to that of Gilmoremys , whereas there are many differences to most species of Pan-Trionychinae. The hyo- and hypoplastron are robust and the lateral and medial processes are short. Moreover, a notch is present at the anterior margin of the holotype, as in Gilmoremys ( Fig. 5 View Figure 5 ). The triangular xiphiplastra might have been sutured to each other along the midline, based on their straight medial margin; however, only two xiphiplastra are preserved in total and the most complete one ( Figs 5G, H View Figure 5 ) has an anteromedial process that possibly prevented such a suture. In the majority of Pan-Trionychinae, on the other hand, the plastron is much more reduced and the lateral and medial processes are longer. Exceptions to this are several species of Nilssonia spp. In those species, the overall plastron morphology is very similar to that of S. baba , including very short lateral and medial processes on the hyo- and hypoplastron and triangular xiphiplastral. The overall plastron morphology thus indicates a closer relationship of Striatochelys either with Plastomenidae or to extant east Asian trionychines.
In conclusion, the carapace of Striatochelys strongly indicates affinities to Pan-Trionychinae. The absence of a preneural is known only in members of this clade, as well as in basal trionychids, and characters indicating a close relationship with Plastomenidae (like the strongly developed ridges on the carapace) can also be found in some juvenile/subadult individuals of Pan-Trionychinae. Additionally, differences in the shapes of costals I and II further support a distinction from Plastomenidae . The plastron of S. baba is very similar to that of Nilssonia spp. and possibly indicates a closer relationship of the former to members of this genus. The very similarly-shaped plastron of the plastomenid Gilmoremys , on the other hand, probably represents a convergence.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |