
Hyalinobatrachium iaspidiense
|
publication ID |
https://doi.org/ 10.5281/zenodo.200895 |
|
DOI |
https://doi.org/10.5281/zenodo.5658433 |
|
persistent identifier |
https://treatment.plazi.org/id/E57687EB-FFB9-5F10-F3A6-E161FCDC1324 |
|
treatment provided by |
Plazi |
|
scientific name |
Hyalinobatrachium iaspidiense |
| status |
|
Hyalinobatrachium iaspidiense View in CoL
( Fig. 5 View FIGURES 5 )
Centrolenella iaspidiensis Ayarzagüena, 1992: 23 View in CoL .
Centrolene iaspidiensis Duellman, 1993: 50 View in CoL .
Hyalinobatrachium iaspidiense Myers and Donnelly, 1997: 16 View in CoL . Hyalinobatrachium nouraguensis Lescure and Marty, 2000: 74 View in CoL , synonym. Hyalinobatrachium nouraguense Kok and Castroviejo-Fisher, 2008: 48 View in CoL .
Type locality. Quebrada de Jaspe, San Ignacio de Yuraní, (04°55’N, 61°05’W; 800–1000 m) Bolívar, Venezuela.
Diagnosis. (1) Dentigerous processes on vomer and vomerine teeth absent; (2) snout truncate in dorsal and lateral view; (3) tympanum covered by skin, not visible through skin; (4) dorsal skin from smooth to shagreened in life and preservative, (5) presence of small cloacal enameled warts; (6) parietal peritoneum transparent, pericardium transparent, visceral and hepatic peritonea white, all other peritonea transparent; (7) liver bulbous; (8) humeral spine absent; (9) webbing formula of fingers III (2– – 2) – (2– – 2+) IV; (10) webbing formula of toes I (1 – 1+) – (2+ – 2 1/3) II (1 – 1 1/3) – (2+ – 2 1/4) III (1 – 1+) – (2 1/2 – 2 3/4) IV (2+ – 2 1/4) – (1+ – 1 1/4) V; (11) enameled ulnar and tarsal folds; (12) nuptial pad Type-V composed by a group of packed glands and situated in the medial, dorso-lateral internal side of Finger I, glands not present in other fingers, prepollex not evident from external view; (13) Finger I longer than Finger II; (14) eye diameter larger than width of disc on Finger III; (15) coloration in life: dorsum light green with big irregular darker green patches, black dots, and minute melanophores, bones white; (16) coloration in preservative: cream with big irregular white patches and black dots; (17) iris yellow with dark grey flecks; (18) minute melanophores not extending throughout fingers and toes except base of Finger IV and Toe V; in life, tip of fingers and toes white; (19) advertisement call composed by a single pulsed note lasting 0.05– 0.10 s, dominant frequency of 4220.5–5000.5 Hz, males call from the underside of leaves; (20) fighting behavior unknown; (21) egg clutches deposited on the underside of leaves, males often on the same leaf than eggs; (22) tadpole not described; (23) medium size adult males, SVL = 18.7–22.2 (20.3 ± 0.8, N = 25) mm and one female 22.0 mm.
Comparisons. The following unique combination of phenotypic characters differentiates Hyalinobatrachium iaspidiense from all other species in the genus: snout truncated in dorsal and lateral views, tympanic membrane and annulus not appreciable in life, pericardium transparent, hand webbing formula III (2- -2) – (2–-2+) IV, dorsal coloration in life light green with big irregular darker green patches and black dots, dorsal coloration in preservative cream with big irregular white patches and black dots, iris coloration in life yellowish, clearing out towards eyelids, with dark flecks, coloration of bones in life white, coloration of hands and feet in life white, and a pulsed single note advertisement call without frequency or amplitude modulation, lasting 0.05– 0.10 s and with dominant frequency of 4220.5–5000.5 Hz.
Morphological, bioacoustic and genetic evidences allowing the differentiation between Hyalinobatrachium iaspidiense and all other species of Hyalinobatrachium from the GS are summarized in Figures 2 View FIGURES 2 , 3, 5 and Tables 1, 3.
Remarks. To address the question of whether or not there are morphological differences between Hyalinobatrachium iaspidiense and H. nouraguense , we compared the type material of both species plus additional specimens from Guyana (N = 8), French Guiana (N = 4), Suriname (N = 11), and Venezuela (N = 2). We could not find any difference in the studied characters. Furthermore, we found descriptive lapses in both Lescure and Marty (2000) and Cisneros-Heredia and McDiarmid (2007). These are not preservation artifacts as stated by Yáñez- Muñoz et al. (2009). Both works described the pericardium of H. nouraguense as white (versus transparent in H. iaspidiense ); however, all the specimens examined (including the type series of H. nouraguense and other material from French Guiana) have a transparent pericardium ( Fig. 5 View FIGURES 5 D). The morphological characters, therefore, do not allow us to separate two species.
We also compared the morphology of those specimens with that of the holotype of Hyalinobatrachium mesai (the only known specimen for this species). The only consistent difference was that H. mesai has green bones in life and those of H. iaspidiense and H. nouraguense are white (see also the Remarks section for H. mesai ).
Our bioacoustic analyses show, in concordance with the morphological data, that there are no divergent features between the calls of H. iaspidiense , H. mesai and H. nouraguense . The description of the call of H. mesai by Barrio-Amorós and Brewer-Carías (2008) indicates that it is longer than that of H. iaspidiense and H. nouraguense . However, the audiospectogram pictured in their publication has too little resolution to be properly compared. Our own analysis indicates that there are no differences in the calls between H. iaspidiense and H. mesai .
Cocroft et al. (2001) report an unidentified species of Hyalinobatrachium from the Amazonian foothills of the Peruvian Andes in Manu National Park. They provide a photograph and the advertisement call of a single male. The specimen fully corresponds to the description here provided for H. iaspidiense and the parameters and structure of its advertisement call falls within the variability of H. iaspidiense . Rivera and Knell (2006) report a photograph of a specimen from the Amazonian lowlands of Tapiche, Loreto, Peru, which shares all of its morphological characters with H. iaspidiense . Photographs of six specimens collected from Río Ituxi, Amazonas, Brazil fully agree with the morphological characteristics of H. iaspidiense (J.P. Caldwell, unpublished data). The sequences obtained from tissue samples of these Brazilian specimens included in our study further support their identification (Fig. 3) and corroborate the morphological analysis. Accordingly, we assign all these specimens to H. iaspidiense , extending its distribution from its most western record in Brazil and Peru.
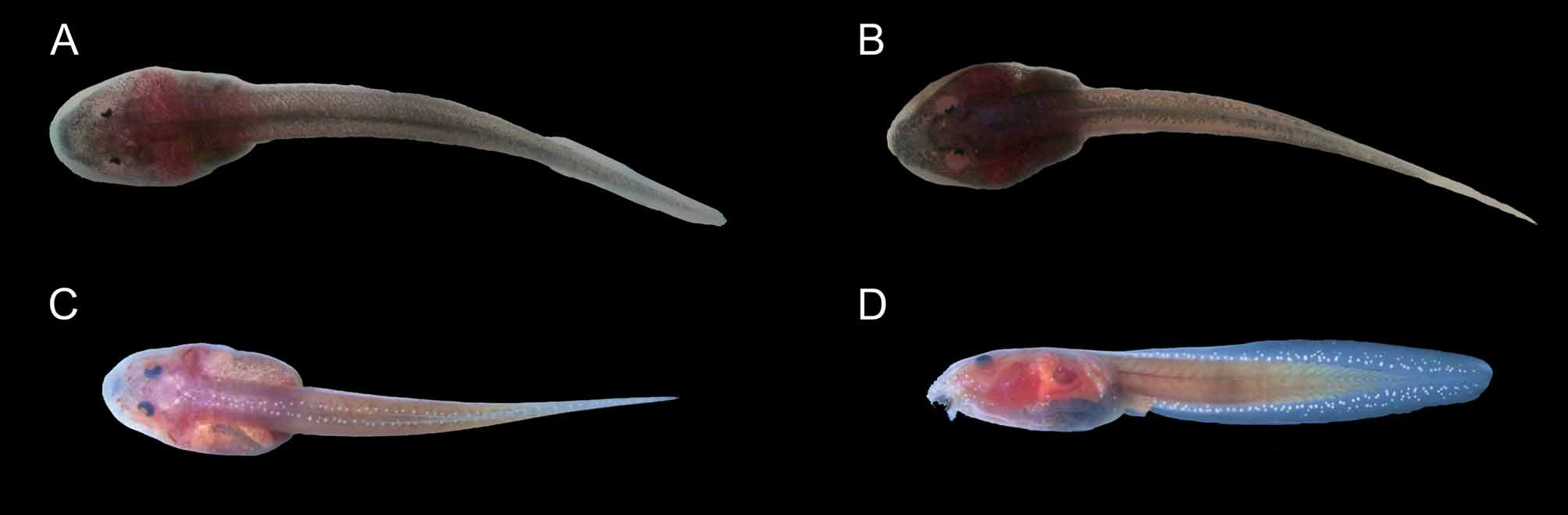
Biology and tadpole. Very little information is available. Position of calling males and advertisement call are described in Señaris and Ayarzagüena (2005) and Lescure and Marty (2000). Here we provide new information from the known locality in Guyana (Ernst et al. 2005, 2006). Calling males were always found on the underside of leaves, usually facing leaf axils. Up to three males were observed calling from the same tree usually during and after rain. Individuals of Hyalinobatrachium iaspidiense occupied lower forest strata (≈ 1.0–2.0 m) in vegetation overhanging fast flowing segments of the creek, as compared to the sympatric H. mondolfii , whose calling sites were located in higher strata (≈ 4.0–6.0 m). Calling males of H. iaspidiense were found near by (≤ 5 cm) clutches containing 22 (in all cases recorded, N = 4) relatively large white eggs. A clutch containing 22 semi-developed tadpoles ( Gosner 1960; stages 21–22) attached to a leaf overhanging a medium sized black water creek was collected on March 15, 2004 (tadpoles deposited at SMNS under field number MABT0104, no collection number assigned), and transferred to a plastic aquarium. The leaf was fastened to the lid of the container, which was filled with 10 cm of creek water. Eleven tadpoles had dropped into the container the following day, nine additional tadpoles followed on March 17. It took an additional day before all remaining tadpoles had left the clutch. The tadpoles of H. iaspidiense are typical exotroph, lotic, fossorial tadpoles (eco-morphological guild after Altig & Johnston 1989) with an elongate, vermiform habitus ( Fig. 6 View FIGURES 6 B). Compared to the larvae of the sympatric H. mondolfii , tadpoles of H. iaspidiense do not exhibit a clearly ovoid body. Rather, the anterior half is markedly triangular in shape, narrowing towards the base of the tail. Tadpoles of H. iaspidiense are always much darker than those of H. mondolfii ( Fig. 6 View FIGURES 6 ).
Ecology and distribution. Hyalinobatrachium iaspidiense inhabits the lowland and upland forests of eastern Guiana Shield (50–1000 m) and western Amazon. It has always been found associated with streams. It is known from Brazil ( Cordeiro-Duarte et al. 2002; Yáñez-Muñoz et al. 2009; Avila-Pires et al. 2010; this work), Ecuador ( Yáñez-Muñoz et al. 2009; Guayasamin & North 2009), French Guiana ( Lescure & Marty 2000; this work), Guyana (Ernst et al. 2005), Peru ( Yáñez-Muñoz et al. 2009; this work), Suriname ( Kok & Castroviejo-Fisher 2008), Venezuela ( Ayarzagüena 1992; Señaris & Ayarzagüena 2005; this work), and expected to occur in the Amazon areas between the Ecuadorian and Peruvian localities and the GS.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hyalinobatrachium iaspidiense
| Castroviejo-Fisher, Santiago, Vilà, Carles, Ayarzagüena, José, Blanc, Michel & Ernst, Raffael 2011 |
Hyalinobatrachium iaspidiense
| Kok 2008: 48 |
| Lescure 2000: 74 |
| Myers 1997: 16 |
Centrolene iaspidiensis
| Duellman 1993: 50 |
Centrolenella iaspidiensis Ayarzagüena, 1992 : 23
| Ayarzaguena 1992: 23 |