
Phasmasaurus, Sadlier & Bauer & Shea & Smith, 2015
|
publication ID |
https://doi.org/ 10.3853/j.2201-4349.67.2015.1649 |
|
persistent identifier |
https://treatment.plazi.org/id/E82C87DD-FFBB-FFC9-060A-73EEC81EFBCA |
|
treatment provided by |
Carolina |
|
scientific name |
Phasmasaurus |
| status |
gen. nov. |
Genus Phasmasaurus gen. nov.
Type species. Leiolopisma tillieri Ineich & Sadlier, 1991 designated hereby.
Diagnosis. Moderately large in size, maximum SVL 61 mm ( maruia ) and 64 mm ( tillieri ), with a moderately elongate body, well developed limbs and digits, and very long tail* (maximum tail length range c. 250–300% of SVL respectively).
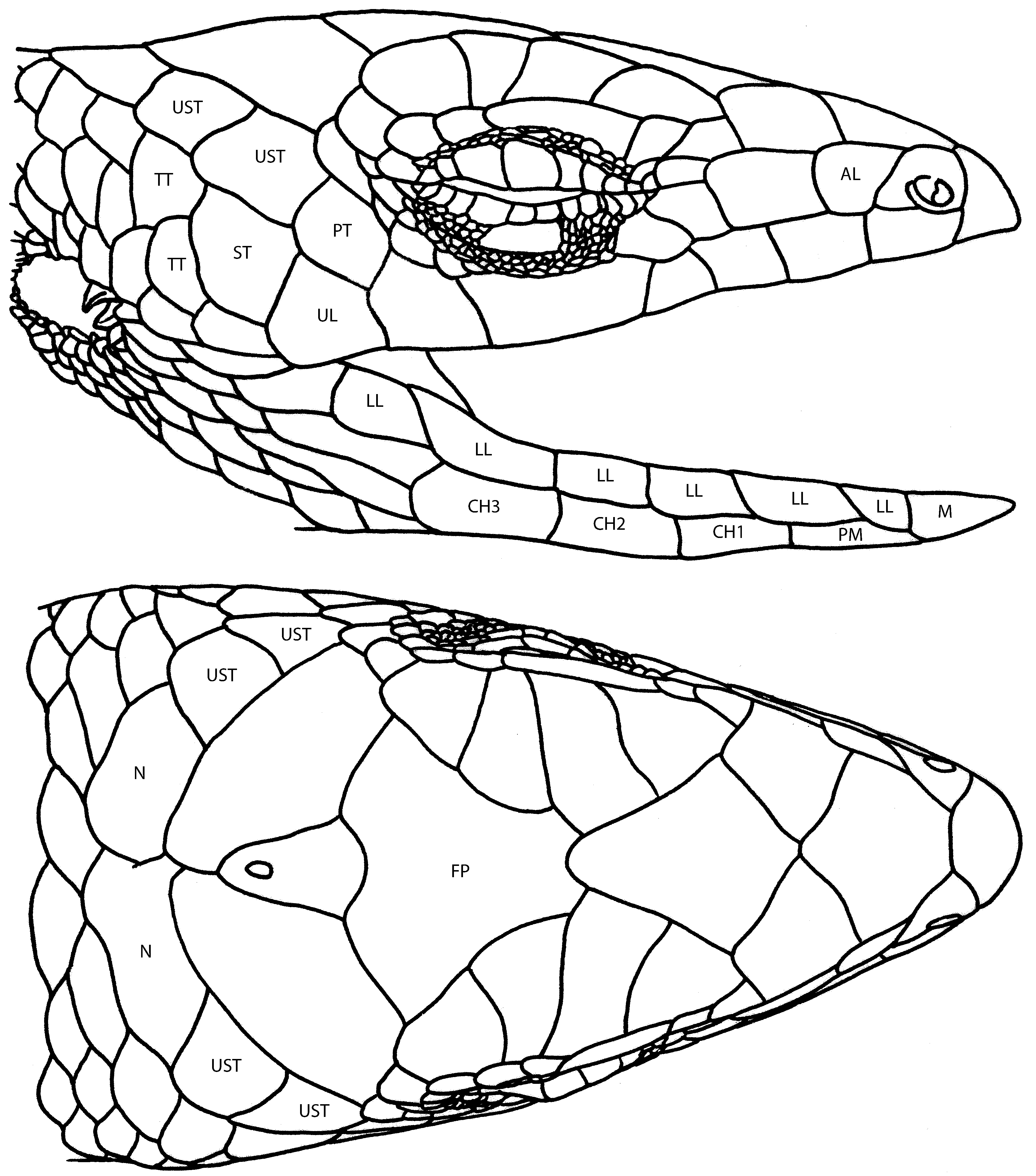
Scalation ( Fig. 5 View Figure 5 ): distinct supranasal absent*; nasal scale lacking a prominent postnasal suture; frontonasal broader than long; prefrontals large; frontal longer than broad; supraoculars four; *frontoparietals fused; interparietal distinct; parietals each bordered by a single nuchal and one ( maruia ) or two ( tillieri ) upper secondary temporal scales; primary temporal single; lower secondary temporal single; tertiary temporals usually two; nasals widely separated; supraciliaries usually seven ( maruia ) or fewer ( tillieri ), reduction through fusion; upper labials usually seven with the fifth subocular and contacting the lower eyelid; postmental contacting first and second lower labial; chinshields three, first pair in broad contact, all in contact laterally with the lower labial scales; *body scales keeled ( maruia three weak keels; tillieri two strong keels).
Osteology: premaxillary teeth 9 ( tillieri ) or 11 ( maruia ); atlantal arches and intercentrum of first cervical vertebrae present as three separate units; 29 presacral vertebrae; *postsacral vertebrae c. 60+; phalangeal formula for the manus of 2.3.4.5.3 and for the pes of 2.3.4.5.4; two pairs of mesosternal ribs.
Reproduction mode: variable within the genus with one species oviparous (egg laying: maruia ) and one viviparous (live-bearing: tillieri ).
The suite of apomorphic character states identified above is modest, but will distinguish Phasmasaurus gen. nov. from all other genera in the Eugongylus group of skinks, including the new genera described here. The key morphological apomorphy uniting tillieri and maruia as sister taxa, and which serves to distinguish them from as a distinct evolutionary entity from most Eugongylus group genera is the exceptionally long tail of the species of Phasmasaurus , characterized by a high postsacral vertebrae number of 60 or more. This character state is otherwise only seen within the Eugongylus group in some Emoia (species in the samoensis group), which are outside the endemic New Caledonian skink radiation. Phasmasaurus gen. nov. can be distinguished from these Emoia in having fused (vs paired) frontoparietals, keeled (vs smooth) body scales, and fused (vs distinct) supranasal scales, although the evolution of supranasal scales within the Eugongylus group, and the polarity of their presence are open to question ( Sadlier, 2010).
Etymology. From the Greek Phasma , a spectre, in the sense of the insect genus Phasma Lichtenstein , type genus for the family Phasmatidae , and for the order Phasmatodea in general, alluding to the phasmid-like appearance and behaviour of species in the group. Gender of the generic name based on the Greek sauros (masculine).
Intergeneric relationships. Support for the two species included in Phasmasaurus as a genetic lineage in the study by Smith et al. (2007) based on one mitochondrial (ND2) and two nuclear (c- mos & RAG-1) genes was high (BPP 1 but with Bootstrap support <50%), and a similar, but slightly lower level of support (BPP 0.95%, bootstrap 71%) was recovered by Ineich et al. (2014). The two species in Phasmasaurus share only a modest suite of morphological apomorphies that serve to define it, including fused frontoparietal scales, keeled body scales, a windowed lower eyelid and a very long tail, all character states that can be found in some other taxa in the Eugongylus group.Additional support for monophyly of the two species as a lineage (outside of the genetic data) comes from unusual shared attributes in habitat preference and behaviour. The strict habitat preference of both species to maquis shrubland and shared unusual aspects of behaviour ( Sadlier et al., 1998) in combination, lend further support to the genetic data for these taxa comprising an independent evolutionary lineage highly divergent from others in the Eugongylus group of skinks.
The scheme of relationships for the Eugongylus group skinks presented by Smith et al. (2007) placed the species here included in Phasmasaurus (and the New Caledonian species here recognized as Caesoris novaecaledoniae ), outside the well-supported lineage that included nearly all other endemic New Caledonian skinks, and within a cluster that included the New Zealand genera inclusive of the Norfolk/Lord Howe Island Oligosoma lichenigera , although this latter group received no support as a lineage in its own right. A more recent molecular phylogeny of Chapple et al. (2009) which sampled extensively the New Zealand Eugongylus group skinks, and which also included representative taxa from the New Caledonian skink fauna for out-group comparison, retrieved a monophyletic New Caledonian skink group represented by the taxa Caledoniscincus austrocaledonicus , Nannoscincus mariei , Marmorosphax tricolor , and “ Lioscincus ” tillieri that was the sister group to the New Zealand + Lord Howe/ Norfolk Island O. lichenigera , and with tillieri now nested within the (albeit reduced) New Caledonian group. The most recent molecular phylogeny of Ineich et al. (2009), which concentrated primarily on the endemic New Caledonian skink radiation and the genetic relationships of the recently discovered Phoboscincus bocourti , but which also included a range of other Eugonglus group genera in the outgroup, placed the species here included in Phasmasaurus as the sister to all the endemic New Zealand skinks (also including Oligosoma lichenigera ), but with no support.
Recognized species. Two, Phasmasaurus tillieri (Ineich & Sadlier) and Phasmasaurus maruia (Sadlier, Whitaker & Bauer) .
Phasmasaurus tillieri (Ineich & Sadlier)
Synonym. Leiolopisma tillieri Ineich & Sadlier, 1991: 344 .
Distribution. Restricted to the southern ultramafic ranges of New Caledonia.
Comments. Endemic to maquis shrubland habitats.
Phasmasaurus maruia (Sadlier, Whitaker & Bauer)
Synonym. Lioscincus maruia Sadlier, Whitaker & Bauer, 1998 ; 335.
Distribution. East-central region ultramafic ranges and the west-central ultramafic ranges as far north as Plateau de Tia and Massif de Kopéto.
Comments. Restricted to maquis shrubland habitats.
Intrageneric relationships. Two genetic-based studies ( Smith et al., 2007; Ineich et al., 2014) retrieve P. tillieri and P. maruia as highly supported (BPP 1.0) sister taxa. However, there is also substantial genetic and morphological differentiation between the two taxa. Nucleotide sequence divergence between P. tillieri and P. maruia for the mitochondrial gene ND2 was around 20.0%, approaching the level of genetic distance found between some genera of New Caledonian skinks (Sadlier & Bauer, unpublished). The high level of genetic differentiation between the two taxa is complemented by a number of differences in morphology and clearly supports their recognition as independent evolutionary entities. While the two species are similar in overall body form in both having long legs, a long tail and an angular head, there are substantial differences in scalation. In particular, the degree of morphological differentiation seen in tillieri , as expressed in the number of species-specific apomorphies, is exceptional in comparison to most other New Caledonian skinks. In particular the strongly bicarinate scales with the keel interrupting the posterior free edge is a trait unique to the species within the context of the Tasmantis group skinks ( Fig. 6a View Figure 6 ). By comparison maruia is plesiomorphic in its head scalation and the weakly tricarinate body scales ( Fig. 6b View Figure 6 ) of this species likely represent the more plesiomorphic form of this character for the genus (see Sadlier, 2010: appendix 1). The two species also differ markedly in reproductive mode, with P. tillieri giving birth to live young while P. maruia has retained the plesiomorphic egg-laying condition. This is the only genus in the Eugongylus group where both modes of reproduction occur. The extent of genetic and morphological differentiation between P. tillieri and P. maruia raises the question of recognising each as a distinct monophyletic genus. While a convincing stand-alone diagnosis could be made for tillieri , the morphological diagnosis for maruia as a lone entity would be weak and lacking in unique apomorphies to further differentiate it from tillieri . For these reasons this action has not been proposed at this time and their inclusion as sister taxa under Phasmasaurus rests primarily on the implications of the phylogenetic relationships retrieved so far from molecular studies.
In overall ecology the two species share traits in behaviour and a strict habitat preference only rarely seen in other endemic New Caledonian skink genera ( Sadlier et al., 1998). Both are restricted solely to maquis shrubland ( Lacertoides is the only other taxon of skink reliant on this habitat type), with tillieri widespread across the southern ultramafic region and maruia on the adjacent central-west ultramafic ranges. As such, the two species are broadly parapatric, and they act as ecological analogues within their respective regions.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.