
Akodon boliviensis
|
publication ID |
https://doi.org/10.5281/zenodo.293461 |
|
DOI |
https://doi.org/10.5281/zenodo.6196163 |
|
persistent identifier |
https://treatment.plazi.org/id/E91F87ED-FFA9-FF8A-D9DA-8CE2FE8CFDB2 |
|
treatment provided by |
Plazi |
|
scientific name |
Akodon boliviensis |
| status |
|
Akodon boliviensis species group
The great morphological similarity among species of the A. boliviensis group, as well as high levels of intraspecific variability, has precluded a clear and stable taxonomy for the group and in particular for those forms present in northwestern Argentina. This situation, together with the lack of studies of representative series of specimens, including the holotypes and additional specimens collected at the type localities, resulted in the suggestion, along the years, of different taxanomic scenarios, sometimes quite distinct among themselves.
As in previous instances (e.g., Patton et al., 2000), the integration of morphologic and molecular data interpreted in a geographic context has proven usefull to assess species boundaries of a taxonomically confusing group. In this regard, the evidence presented herein indicates the existence of four species of the A. boliviensis group in northwestern Argentina with a new species belonging to this group from central Argentina.
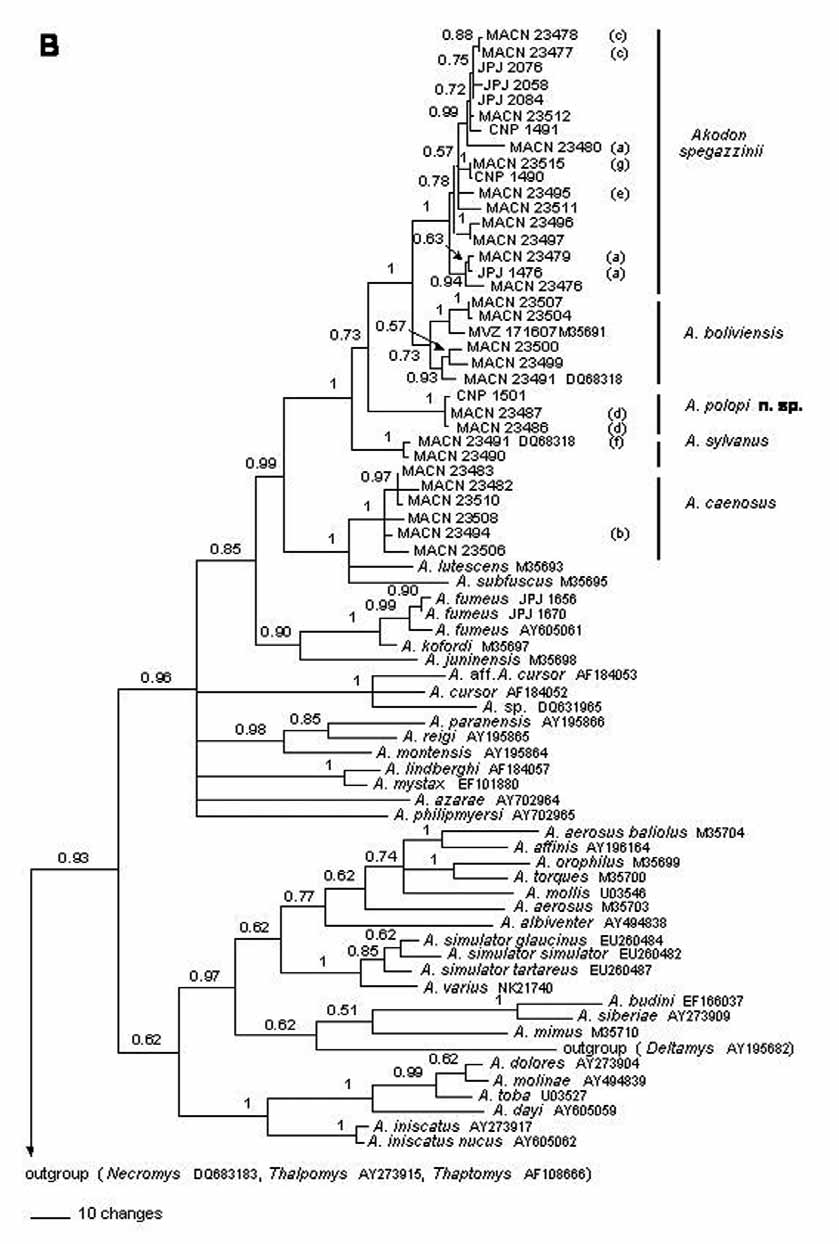
The taxonomic status of A. spegazzinii (including alterus , leucolimnaeus , and tucumanensis ) with respect to A. boliviensis is the most debatable of the taxonomic decisions we make. Based on a small series of specimens, Myers et al. (1990) had earlier proposed the valid status of this form. In line, much of the available evidence here considered suggests that A. spegazzinii represents a distinct species from A. boliviensis . For example, they form reciprocally monophyletic groups ( Fig. 1 View FIGURE 1 ) that are allopatrically distributed; in addition, they are clearly identifiable in morphometric multivariate space and morphological grounds (see comparison above). Moreover, these species have different karyotypes, including the presence of heteromorphism in the largest autosomal pair and the X chromosome in A. spegazzinii ( Gardner & Patton, 1976; Barquez et al., 1980; Myers et al., 1990), and different ecological preferences. Our records indicate disjunct distribution for these forms, with A. boliviensis restricted to high altitudinal grasslands in the extreme north of the region and A. spegazzinii extended over many habitats and large areas in south-central portions of the region. In spite of these differences, the genetic divergence betweem them was the smallest among all species pairs of the A. boliviensis group ( Table 12 View TABLE 12 ). In addition, some overlap in morphological and morphometric characters was evident. Moreover, we could not evaluate the constancy of these differences because we failed to capture both species in sympatry. So, additional studies are needed in the north of the northwestern Argentinean region and in possible areas of sympatry to test the constancy of the observed differences here reported.
The taxonomic status of A. tucumanensis , A. alterus , and A. leucolimnaeus remained controversial despite the pioneering work by Myers et al. (1990). While all these forms were treated as subspecies or allied to A. spegazzinii by some authors (e.g. Myers et al. 1990; Díaz et al. 2000; Pardiñas et al. 2006; Jayat et al., 2008a), others considered them as valid species (e.g. Barquez et al., 1991; Mares et al., 1997; Díaz, 1999; Díaz & Barquez, 2007; but see Galliari et al., 1996 about the status of A. tucumanensis ). In spite of the extended usage of the specific epithet alterus in previous works (e.g. Capllonch et al. 1997; Díaz et al. 1997; Mares et al. 1997; Díaz 1999; Díaz & Barquez 2007) none of these references was based on a detailed study of extensive topotypical series nor the use of molecular characters. Blaustein et al. (1992) were the only authors who tested the status of alterus (with respect to tucumanensis ). However, they erroneously assumed that specimens from El Infiernillo, Tucumán, belonged to alterus ; then, they did not truly test the taxonomic status of A. alterus . They also failed to separate these specimens from A. tucumanensis from low altitude Yungas forest of Tucumán. In words of Blaustein et al. (1992, pag. 11) “... we believe that any conclusion on the taxonomic status of the two forms is untimely.” The status of A. leucolimnaeus was even more dubious and, until the preliminary observations of Galliari et al. (1996), it was considered as a synonym of Necromys lactens ( Cabrera, 1961; Reig, 1978, 1987; Mares et al., 1997). However, Galliari et al. (1996) ranked leucolimnaeus as a valid species of Akodon (see also Musser & Carleton [2005] and Pardiñas et al. [2006]), view that contrasts with our results. All the available evidence indicates that the nominal forms alterus , leucolimnaeus , and tucumanensis must be considered junior synonyms of A. spegazzinii . Haplotypes from specimens coming from the type localities of these fall within the spegazzinii clade ( Fig. 1 View FIGURE 1 ). The genetic divergence among haplotypes of this clade is low (1.2% in average) and we found no clear morphological or morphometric differences among these forms. Most of the observed differences in coloration among populations may relate to the great environmental heterogeneity inhabited by A. spegazzinii ( Fig. 10 View FIGURE 10 ).
In the A. boliviensis clade, two haplotypes recovered from specimens from northern Salta were more closely related to the haplotype of a Peruvian specimen (MVZ 171607) than to those of other three Argentinean specimens ( Fig. 1 View FIGURE 1 ). This phylogeographic break, together with some morphological, morphometric and genetic differences between these two groups also suggest the possiblity of unrecognized biological diversity within this group. This scenario is similar to that observed in N. amoenus , another highland inhabitant (D’Elía et al., 2008). Notwithstanding, the studied material of A. boliviensis is scarce and the difference between both clades is moderate (2.1% of average genetic divergence), so we point that further research is needed on this area.
Myers et al. (1990) considered A. caenosus as a subspecies of A. lutescens (as A. puer caenosus in that publication). They highlighted subtle morphometric, morphologic, and cytogenetic characteristics that distinguished specimens from northwestern Argentina from those of Peru and Bolivia. Our phylogenetic analyses include haplotypes recovered from Argentinean specimens assignable to caenosus and a Peruvian specimen belonging to lutescens . These haplotypes form a paraphyletic group respect to A. subfuscus ( Fig. 1 View FIGURE 1 ). This topology, if lutescens and subfuscus are not to be considered conspecifics, suggests a specific status for A. caenosus . Moreover, the average genetic distance between the Peruvian haplotype of lutescens (MVZ 171612) and those from northwestern Argentinean specimens of A. caenosus is relatively high (3.5%); the average divergence value among specimens of A. caenosus is much lower (1%). Although, there are not clear morphological and morphometric differences among our material of A. caenosus and the values offered by Myers et al. (1990) for A. lutescens , the alternative position of considering A. subfuscus and A. lutescens conspecific seems less supported. These species differ by a substantially high (5.1%) average genetic distance. Furthermore Myers et al. (1990) detailed several morphological characters that distinguish A. lutescens from A. subfuscus .
Although we failed to obtain small specimens of Akodon from the type locality of A. aliquantulus (we studied ca. 100 individuals of Akodon from this locality and its surroundings and more than 390 from Tucumán province), its diagnosis does not permit us to distinguish the two type specimens from A. caenosus . Moreover, the haplotypes of three small Akodon sequenced from Catamarca and Tucumán were remarkably similar to haplotypes of A. caenosus topotypes (with average genetic distance lower than 1%). On these bases (see also our Species Account discussion above) we place aliquantulus under the synonymy of A. caenosus . This study corroborated the proposition of Jayat et al. (2007a) of placing A. sylvanus as a valid species of the A. boliviensis group. In the MP analysis A. sylvanus appears as the sister species of A. polopi ; in accordance to low support of this clade (JK<50) it is not recovered in the BA where A. polopi is sister to the A. boliviensis - A. spegazzinii clade.
Akodon polopi , the new species here described, is a clearly distinct member of the A. boliviensis group, with several diagnostic characters coming from different sources of evidence; not surprinsingly, it was early mentioned as a possible new subspecies by Elio Massoia or a new species (see Polop, 1989). This species, known only from high altitude grasslands of central Argentina, is the only species of the A. boliviensis group inhabiting the Sierras Grandes range, a medium altitude mountain system isolated (ca., 600 km) from the main Andean chain by low elevation arid and semiarid environments. Data at hand are insufficient to pose a biogeographic scenario accounting for the diversification of A. polopi and the remainder species of the A. boliviensis group. Notwithstanding, Pampa de Achala has been mentioned as a biogeographic refuge and an area of endemism for other taxa ( Polop, 1989, and references there).
Relationships among species of the Akodon boliviensis group are well resolved and mostly coungrent among analyses. Differences relate to the position of A. polopi and A. sylvanus .
The present study, as several previous ones, reaffirms the phylogenetic legitimacy of both the boliviensis group and extends our knowledge of its contents, limits, and geographic ranges of its member species. However, we assert that additional research is still needed before an adequate picture of Andean Akodon can be acquired. In this line, more field work coupled with the study of specimen series housed in research institutions is essential.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.