
Trichocolletes pulcherrimus Michener, 1965
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.64.2012.1589 |
|
publication LSID |
lsid:zoobank.org:pub:42F22568-9A99-4F96-A422-F020AA558F90 |
|
persistent identifier |
https://treatment.plazi.org/id/E93C9B69-4641-5D4E-5DB0-FAF7FC767F1D |
|
treatment provided by |
Carolina |
|
scientific name |
Trichocolletes pulcherrimus Michener |
| status |
|
Trichocolletes pulcherrimus Michener View in CoL
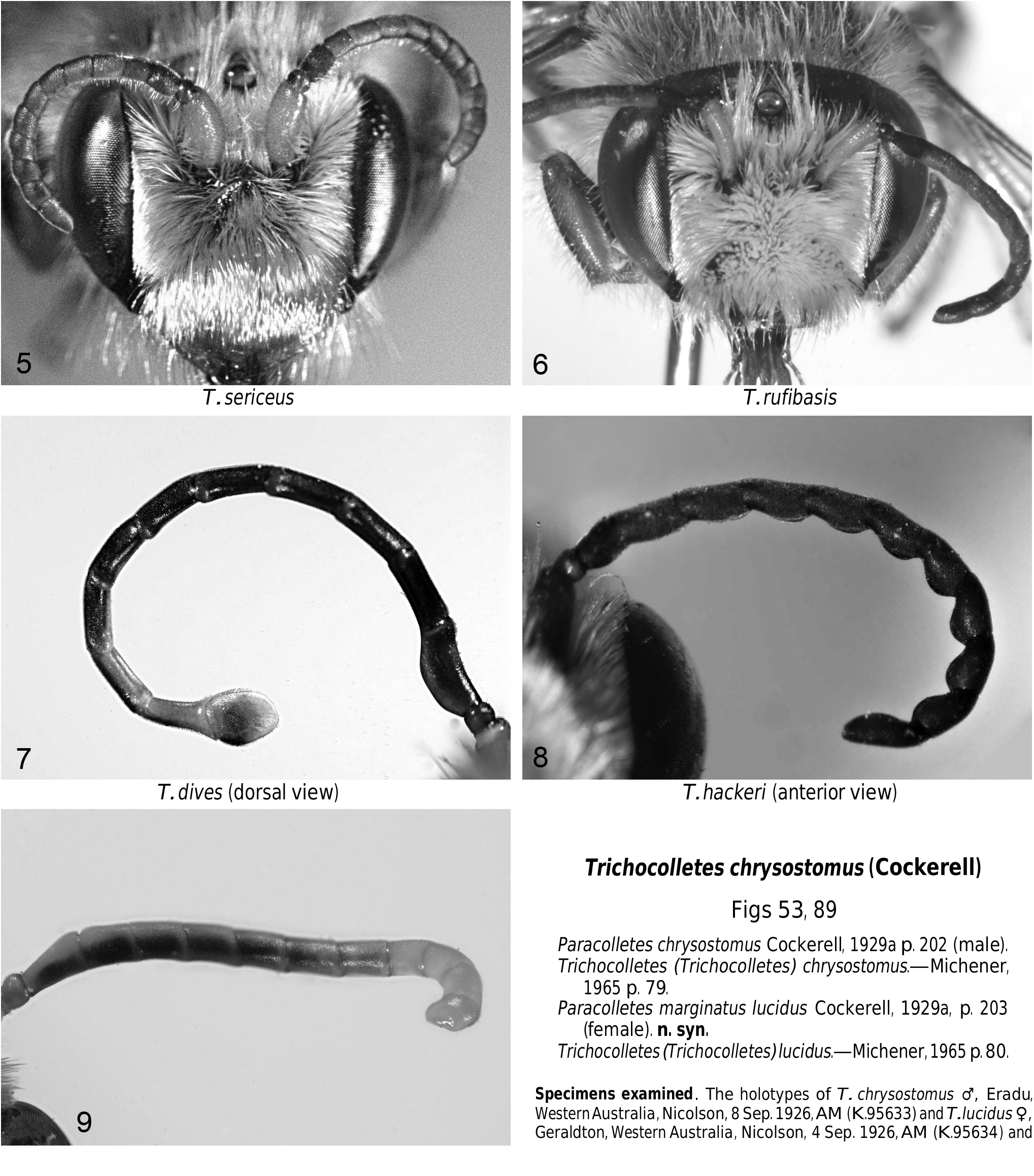
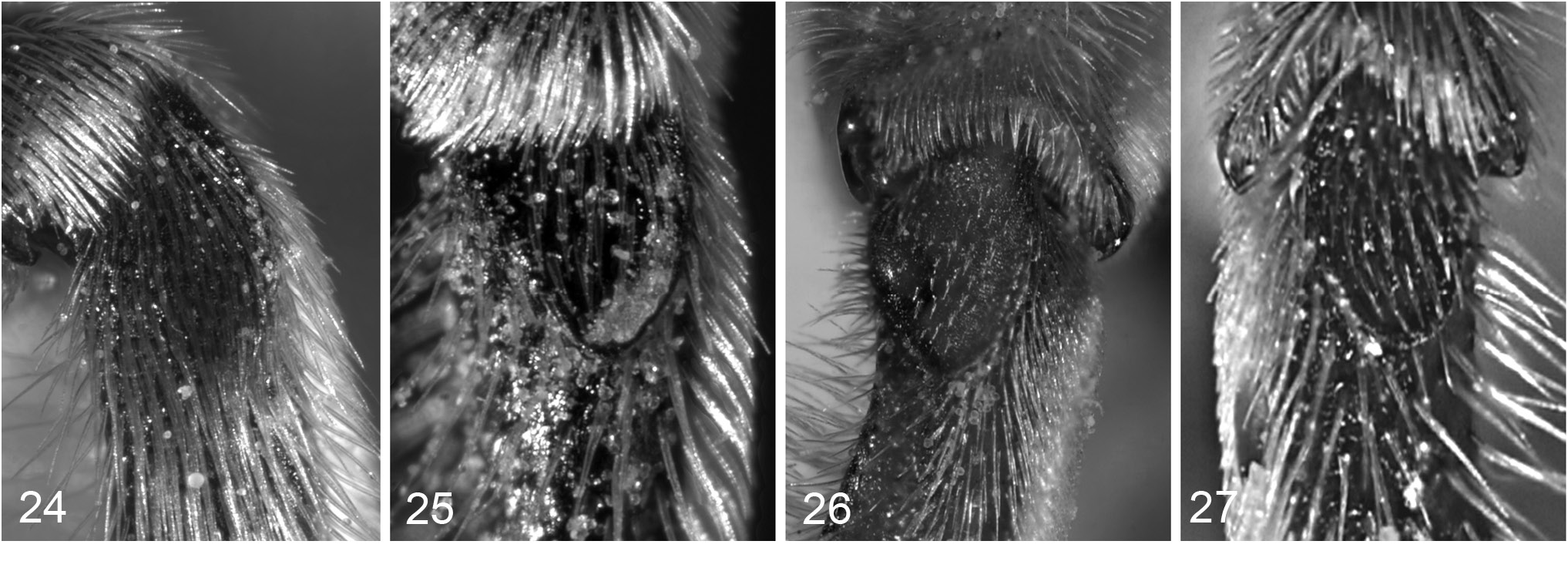
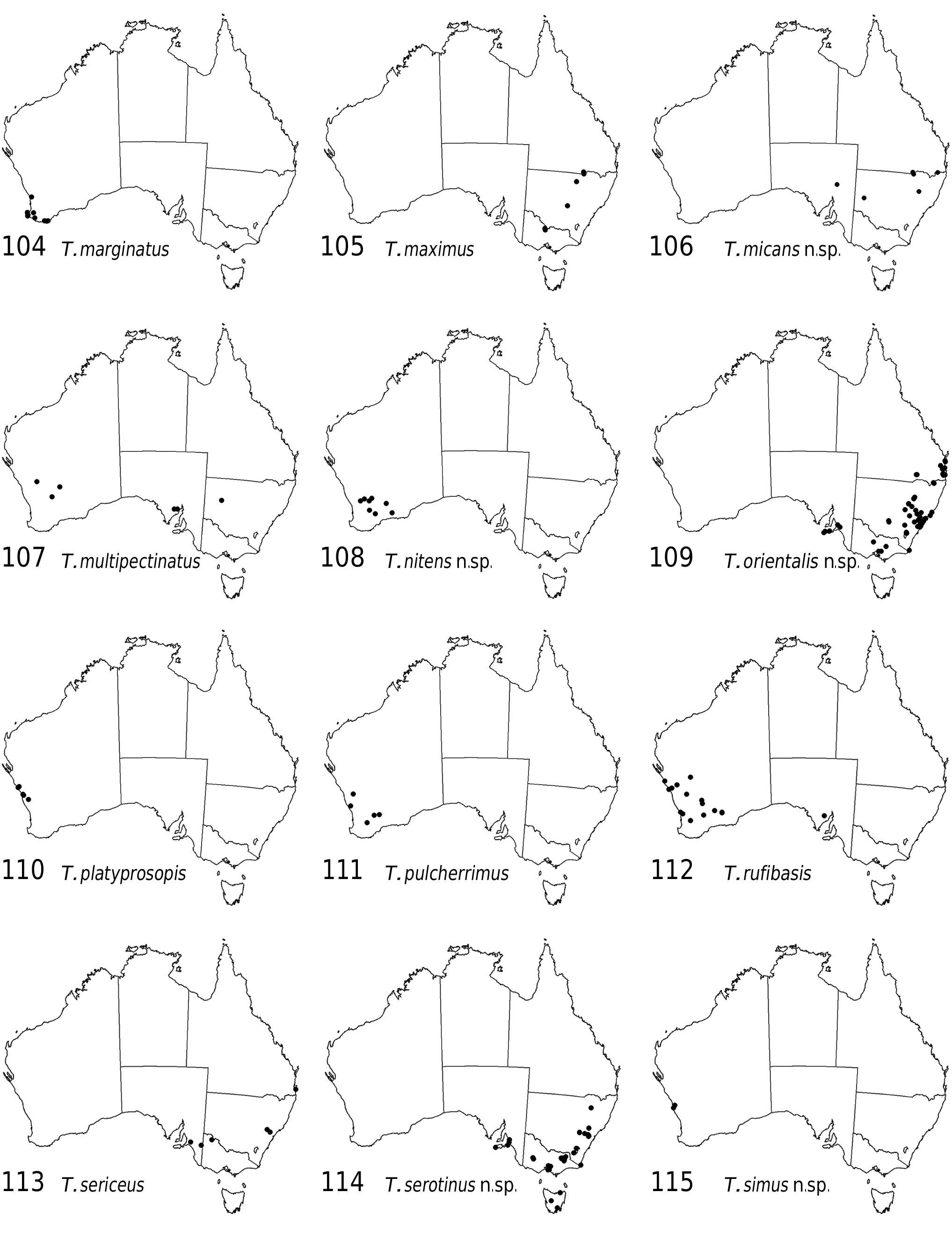
Figs 9 View Figs 5–9 , 22 View Figs 20–23 , 26 View Figs 24–27 , 111 View Figs 104–115
Trichocolletes (Callocolletes) pulcherrimus Michener, 1965, p. 265 View in CoL .
Specimens examined. Holotype ♂, Narrogin, Western Australia, 1 Nov. 1935, WAM (65–727) and the following. Western Australia: ♀, Dumbleyung, 14 Nov. 1966, H. Udell, WAM (9007); 4♀, 75 km E Hyden, 24–27 Oct. 1985, T. F. Houston, on flowers of Gastrolobium spinosum, WAM View in CoL (8997–9000); 2♂, 61 km E Hyden, 14 Oct. 1979, T. F. Houston, on flowers of Burtonia hendersoni, WAM (9001–02); ♀, Regans Ford Moore River , 11 Nov. 1976, S. M. Postmus, on Jacksonia sp., WAM (9006); ♀, 10 S Perenjori, 1 Nov. 1958, E. F. Riek, ANIC; 3♀, 19 km ESE Southern Cross, 28 Oct.1978, T. F.Houston, on flowers of Gompholobium viscidulum, WAM View in CoL (9003–05).
Diagnosis
Length 16–18 mm; eyes not hairy; metasoma orangebrown with indistinct metasomal bands; flagellum banded dorsally in both sexes. Male hind tibia strongly expanded, without spurs; apical segment of flagellum truncate, slightly expanded on one side; fore trochanter with large spine. Female scape and more than half clypeus orange-brown.
Remarks. Detailed descriptions can be found in Michener, 1965.
In 1965, Michener proposed division of the genus Trichocolletes into two subgenera, with this species as the sole member of the subgenus Callocolletes . At that time, the Colletidae family was regarded as a primitive lineage, but molecular evidence now indicates that it is one of the more derived (Danforth et al., 2006) and that separation of Trichocolletes from other Australian colletids occurred relatively early (Almeida and Danforth, 2009). However, the premise on which the subgenus was based remains unaltered. If the hind basitibial area of T. pulcherrimus females is intermediate in nature between those found in the remaining Trichocolletes species and those of other colletids, it might indicate that T. pulcherrimus occupies a basal position within the genus.
The use of molecular evidence from fresh material to test this hypothesis was, unfortunately, not possible as attempts to recollect specimens were unsuccessful, but the increased number of species now available made possible reassessment of the morphological evidence. Although the male exhibits a number of striking modifications including the absence of hind tibial spurs, Michener (1965, 2007) chose to give greater weight to the hind basitibial area and details of wing venation.
The basitibial area of females is difficult to observe because of a dense cover of stiff setae. When the setae are removed, the contiguous depressions at the base of each seta form a coarsely areolate surface ( Fig. 22 View Figs 20–23 ). Interpretation of the resulting structure can be difficult. Frequently specimens are found with the setae worn down to the tops of the depressions making the actual surface difficult to detect and producing an apparent rim to the basitibial area. A second difficulty is that even when setae are removed cleanly, the edge of the basitibial area may be coarsely serrated where it intersects the depressions (as shown for T. brachytomus n.sp. in Fig. 21 View Figs 20–23 ). It is unclear whether or not this state should be classified as carinate.
A better character to use may be the hind basitibial area of males. Comparison of the basitibial areas of males of eight species with those of conspecific females confirmed that the degree of elevation of the basal area from the surrounding surface of the tibia in males reflected that observed for the females after removal of the setae. Of the
T. maximus T. brachytomus n.sp.
T. pulcherrimus T. lacaris n.sp. 38 species for which males are known, five, T. brachytomus n.sp., T. dives , T. lacaris n.sp., T. pulcherrimus and T. tuberatus n.sp., were found to have basitibial areas at least as well developed as in T. pulcherrimus ( Fig. 26 View Figs 24–27 ).
The second criterion used by Michener (1965) was the ratio of the lengths of the pterostigma and prestigma. This measurement is difficult to make accurately and involves a degree of subjectivity. It was found ( Table 1, Fig. 28 View Fig ) that the ratio for T. pulcherrimus lies in the middle of the range for all species. Using 10 specimens each of T. orientalis n.sp. and T. venustus , it was estimated that the standard deviation of the measurements was 0.1.
Trichocolletes pulcherrimus males are unique in having a combination of unusual characters, but with the exception of the loss of the hind tibial spurs, similar characters occur separately or in other combinations in a number of species: projections are found on the fore trochanters of males of T. avialis n.sp. and T. nitens n.sp.; modified legs are found in T. aureotinctus , T. avialis n.sp., T. burnsi , T. dives , T. dowerinensis , T. hackeri and T. macrognathus n.sp.; and modified flagella occur in T. dives and T. gelasinus n.sp.. These male characters are not correlated with the female characters discussed above and it is concluded that the evidence presently available does not support recognition of the subgenus Callocolletes .
Distribution. Southwestern Australia (AW, MAL, SWA) ( Fig. 111 View Figs 104–115 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trichocolletes pulcherrimus Michener
| Batley, Michael 2012 |
Trichocolletes (Callocolletes) pulcherrimus
| Michener, C 1965: 265 |