
Krabimeryx Métais, Chaimanee, Jaeger, and Ducroq
|
publication ID |
https://doi.org/10.1038/s41598-021-96221-x |
|
DOI |
https://doi.org/10.5281/zenodo.5645755 |
|
persistent identifier |
https://treatment.plazi.org/id/E97287E4-4C42-7A0C-1FEA-FB6CFADACDC2 |
|
treatment provided by |
Valdenar |
|
scientific name |
Krabimeryx Métais, Chaimanee, Jaeger, and Ducroq |
| status |
|
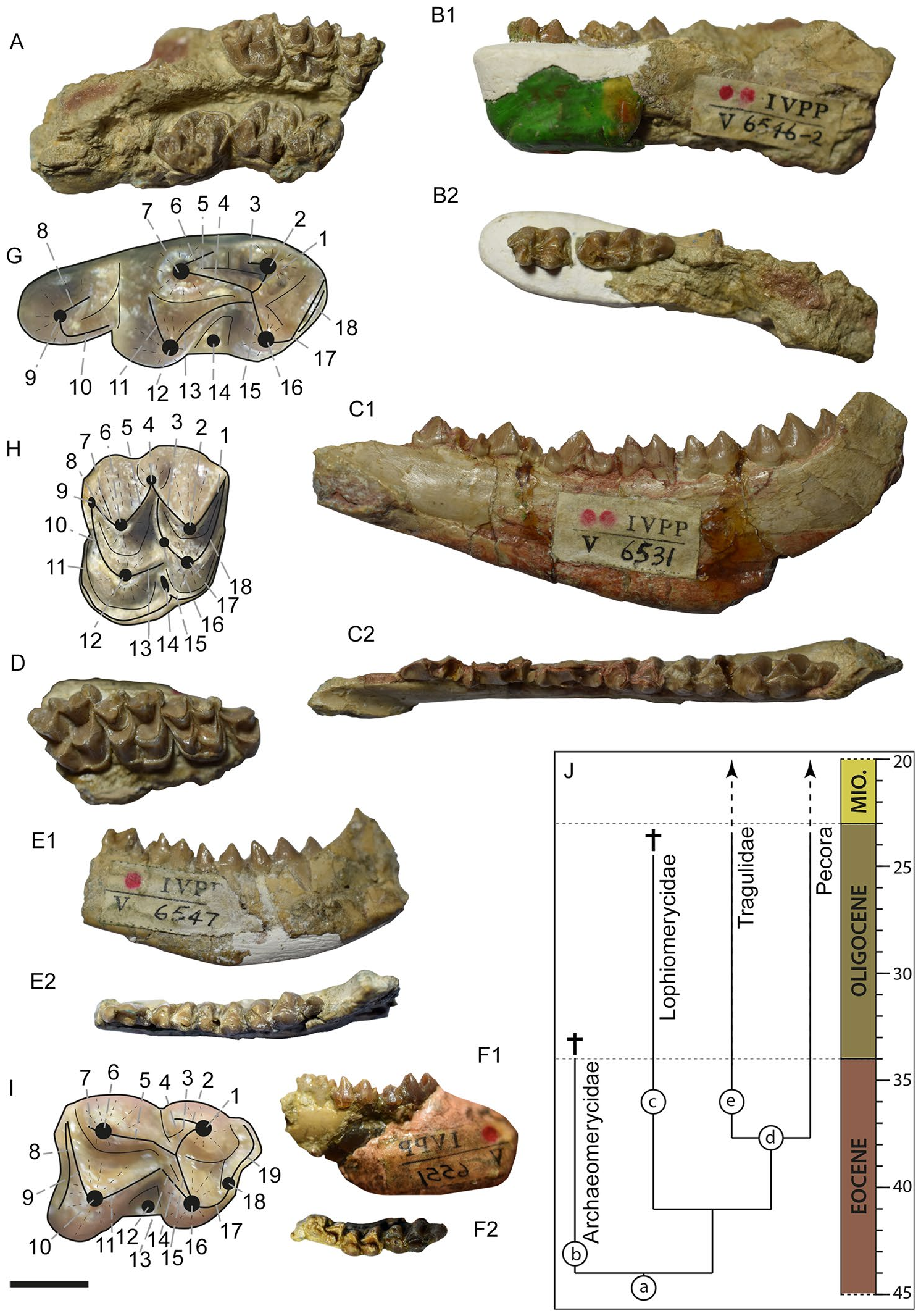
Genus Krabimeryx Métais, Chaimanee, Jaeger, and Ducroq , 200117.
Etymology. Krabi— from Krabi Basin, where the fossils were found, and— meryx is the Greek word for ruminant.
Diagnosis [modified after Métais et al. 17]. Small primitive ruminant with lower molars morphologically close to those of Zhailimeryx . Krabimeryx differs from Zhailimeryx in: more laterally compressed lingual cuspids in the lower molars; an entoconid displaced to anterior with respect to the hypoconid; the lack of both a paraconid and a hypoconulid in m1 and m2; a p4 with a mesolingual conid that is located more posterior and less individualized; a p4 without a distinct posterolingual conid. Krabimeryx differs from Lophiomeryx by less selenodont labial cuspids in the lower molars, the presence of a developed external postmetacristid, and by a distinct groove on the anterior side of the entoconid, the entoconidian groove. Krabimeryx can be distinguished from Iberomeryx in having a well-marked entoconidian groove; the lack of a clear external postprotocristid; the third lobe of m3 not forming a complete buckle; and a more transversely compressed hypoconulid in the m3. Krabimeryx possesses a huge notch in lingual view between the entoconid and the third lobe in the m3.
Type species. Krabimeryx primitivus Métais, Chaimanne, Jaeger, and Ducroq, 200117.
Included species. Krabimeryx gracilis nov. comb. (Miao, 198220).
Krabimeryx gracilis nov. comb. (Miao, 198220).
Figure 1A View Figure 1 and Figure S1 View Figure 1 .
*v pars1982 Lophiomeryx gracilis — Miao: 532, Table 3, Figs. 6 and 720.
v non1982 Lophiomeryx gracilis ?— Miao: 536, Fig. 820.
v pars1987 L. gracilis — Janis: 21133.
v pars 1997 L. gracilis — Vislobokova: Fig. 321.
v pars 2000 L. gracilis — Guo, Dawson, and Beard: 247, Table 214.
v pars 2001 L. gracilis — Métais, Chaimanee, Jaeger, and Ducroq: 239, 24117.
v pars 2012 L. gracilis — Mennecart: 6234.
Scientific Reports |
3
Neodiagnosis. Krabimeryx gracilis has an m2 that is wider than the m3; this is the other way round in K. primitivus. Moreover, the entoconid is less anterior relative to the hypoconid in K. gracilis than it is in K primitivus. Te ectostylid is large in K. gracilis , while it is absent in K. primitivus. Te cingulum on the upper molars in K. gracilis is more developed than in K. primitivus.
Holotype. IVPP V 6546 , partial skull with right and lef M1–M3 ( IVPP V 6546-1 ) and an associated right fragmented mandible with m2–m3 ( IVPP V 6546-2 ) found in occlusion with the skull.
Additional material. IVPP V 6549 , right m3 on fragmented mandible; IVPP V 6550 lef fragmented mandible with m1–m2; IVPP V 26638 View Materials , right m1. Measurements are given in Table S1.
Localities. Shinao Basin , Panxian County, Southwestern Guizhou, China; Xiaerhete locality , Jiminay County, Xingjiang , China. Late Eocene.
Taxonomical attribution. Te herein described specimens were first attributed to the genus Lophiomeryx 20. However, the thorough reassessment of the specimens now leads to the conclusion that Lophiomeryx gracilis sensuMiao 20 contains three different species and genera, but none of them can be assigned to Lophiomeryx .
Based on the presence of a strong lingual cingulum in upper molars and a short anteroposteriorly oriented postprotocrista, as well as the absence of a premetacristid and an anterior fossa widely open in the lower molars, we can conclude that the specimens, IVPP V 6546-1, IVPP V 6546-2, IVPP V 6549, and IVPP V 6550, belong to Lophiomerycidae or Tragulidae 35, 36. However, the absence of a large paraconid and the absence of an elongated external postmetacristid distinguish the specimens from primitive Tragulidae 17, 36. In Zhailimeryx jingweni, the cuspids are more slender than in the herein described specimens 14, a feature the taxon shares with K. primitivus. In Z. jingweni, m1 and m2 are of relative similar width 14, while in K. primitivus and the herein described specimens from Shinao the m2 is clearly bigger than the m 117. Similarly to K. primitivus, the herein described specimens differ from Z. jingweni in its lower molar lingual cusps being more laterally compressed, and in an entoconid that is slightly shifed to anterior with respect to the hypoconid, while it is more posterior in Z. jingweni 14, 17. Furthermore, K. primitivus and the herein described specimens from Shinao both lack the rudimentary paraconid present in Z. jingweni 14, 17.
Like K. primitivus, the here-described specimens differ from Chiyoumeryx nov. gen. (described below) and the Lophiomeryx species L. mouchelini, L. chalaniati and L. angarae by having more massive and more bunomorph lowermolars 16, 17, 24, 34, 37. Furthermore, Zhailimeryx jingweni, K. primitivus, and the herein described specimens differ from Lophiomeryx by the presence of a developed external postmetacristid and by a distinct entoconidian groove on the anterior side of the compressed entoconid 14, 17. In Lophiomeryx , the back fossa of m3 is widely open due to the strong reduction of the posthypoconulidcristid 34, 37. In contrast to this, Krabimeryx primitivus possesses a clearly developed posthypoconulidcristid forming a buckle on the m3 back basin 17, similarly to the specimens from Shinao described here.
Summing up, the general morphology of the teeth in the herein described specimens is most similar to the one observed in K. primitivus. Tey both share a similar huge notch in lateral view between the third lobe of m3 and the entoconid and the entoconidian groove, features that clearly distinguishing them both from Lophiomeryx and Zhailimeryx . Tus, we attribute the specimens IVPP V 6546-1, IVPP V 6546-2, IVPP V 6549, and IVPP V 6550 to the genus Krabimeryx . However, significant differences occur with the type species, ruling out the synonymisation of K. gracilis nov. comb. and Krabimeryx primitivus. While both species are very similar in size, K. primitivus has an m3 wider than m2, while it is the converse for K. gracilis nov. comb. Moreover, the entoconid is less shifed to the anterior with respect to the hypoconid in K. gracilis nov. comb. than in K primitivus. Tere is no ectostylid in K. primitivus, while it is large in K. gracilis nov. comb., forming a transverse cristid between the protoconid and the hypoconid. Te cingulum on the upper molars is more developed in K. gracilis nov. comb. than in K. primitivus.
Due to these differences we decided to create the new combination Krabimeryx gracilis nov. comb.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Ruminantia |
|
InfraOrder |
Tragulina |
|
Family |
Krabimeryx Métais, Chaimanee, Jaeger, and Ducroq
| Mennecart, Bastien, Aiglstorfer, Manuela, Li, Yikun, Li, Chunxiao & Wang, ShiQi 2021 |
L. gracilis
| Mennecart 2012: 62 |
L. gracilis
| Métais, Chaimanee, Jaeger, and Ducroq 2001: 239 |
L. gracilis
| Guo, Dawson, and Beard 2000: 247 |
L. gracilis
| Janis 1987: 211 |
Lophiomeryx gracilis
| Miao 1982: 532 |
Lophiomeryx gracilis
| Miao 1982: 536 |
L. gracilis
| Vislobokova: Fig. 321 . |