
Unesconia, Anker, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4731.1.8 |
|
publication LSID |
lsid:zoobank.org:pub:7E0728D4-2F47-473A-B001-49084FDB7BA3 |
|
persistent identifier |
https://treatment.plazi.org/id/EA68F531-5C60-4B73-E382-60EC7D257AB4 |
|
treatment provided by |
Plazi |
|
scientific name |
Unesconia |
| status |
gen. nov. |
Genus Unesconia View in CoL gen. nov.
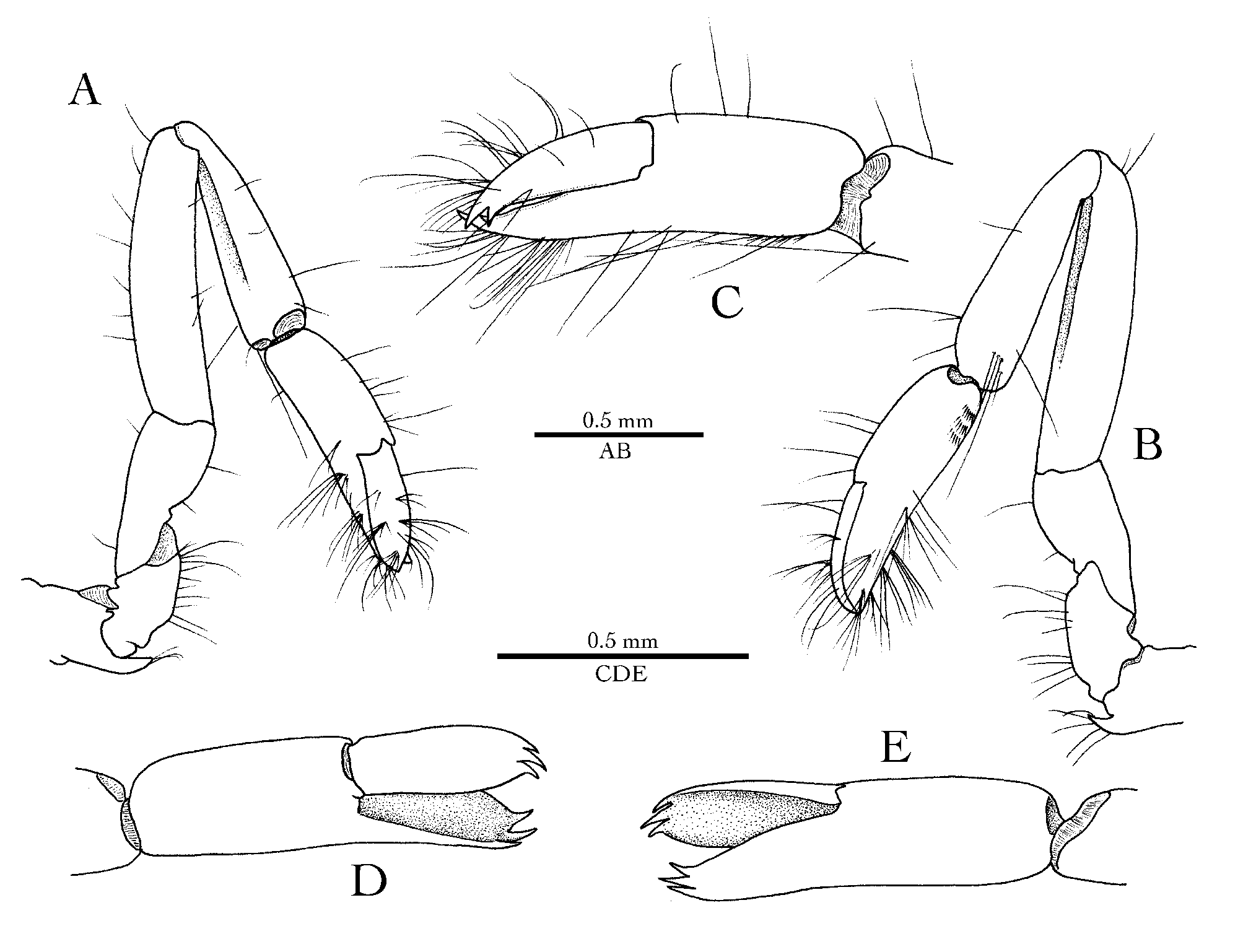
Diagnosis. Small-sized palaemonid shrimp with dorsoventrally flattened body. Carapace smooth, glabrous. Rostrum well developed, dorsally carinate and dentate, ventrally unarmed, lateral carina proximally expanded, armed with strong supraorbital teeth. Orbit well developed, inferior orbital angle distinct; epigastric and hepatic teeth absent; antennal tooth strong, acute. Fourth thoracic sternite without median process. Pleon smooth, glabrous; posteroventral margin of first four pleura rounded, that of fifth pleuron angular, that of sixth pleonite subacutely produced. Telson with two pairs of stout cuspidate setae on dorsal surface and three pairs of spiniform setae on posterior margin. Eyes with cornea well-developed, globular. Antennule with first article ending in stout distolateral tooth; stylocerite small, acute; lateral flagellum with short accessory ramus. Antennal scaphocerite with broad blade and strong distolateral tooth. Epistome unarmed. Mouthparts of non-filtering type. Mandible without palp; incisor process with at least two distal teeth. Maxillule with laciniae broad; palp bilobed distally, ventral lobe with hook-shaped seta. Maxilla with palp (endopod) moderately broad; endite with deep cleft; scaphognathite fairly broad. All maxillipeds with well-developed exopods. First maxilliped with palp (endopod) narrow, short; endite broad; exopod with welldeveloped caridean lobe; epipod moderately developed, bilobed. Second maxilliped of typical palaemonid type; epipod moderately developed, subquadrate, without podobranch. Third maxilliped pediform; coxal lateral plate broadly rounded; basis and ischiomerus distinct, not completely fused; arthrobranch absent. First pereiopods relatively short, stout, fingers subspatulate, with tridentate tips. Both second pereiopods (chelipeds) enlarged, robust; chelae unequal in size, asymmetrical in shape; major chela moderately swollen, bearing two opposing deep fossae, one on pollex (fixed finger) and one on dactylus; minor chela smaller, slenderer, cutting edges of fingers unarmed. Third to fifth (ambulatory) pereiopods relatively slender; ischium, merus and carpus unarmed; propodus armed with few small spiniform setae on ventral margin; dactylus stout, compressed; unguis slender, acute; corpus with one large distal tooth, stouter than unguis, and two or three smaller articulated spinules on ventral margin. Male second pleopod with well-developed appendix masculina. Uropod with lateral lobe of protopod acutely produced; exopod with small distolateral tooth; diaeresis abruptly curved near its lateral end, with four or five strong spiniform setae, latter not extending to exopod’s lateral margin. Gill/exopod formula: 5 pleurobranchs (P1–5), 2 epipods (Mxp1–2), 0 arthrobranchs (Mxp3), 0 podobranchs (Mxp2), 3 exopods (Mxp1–3).
Type species. Unesconia coibensis View in CoL gen. et sp. nov., by original designation.
Other species included. None.
Distribution. Tropical eastern Pacific: presently known only from the Coiba Archipelago, Panama.
Etymology. The new genus is named in honour of the United Nations Educational, Scientific and Cultural Organization (UNESCO), for declaring the Coiba National Park, where the type locality of the new genus’ type species is situated, a UNESCO World Heritage Site, along with many other important protected areas throughout the world. The name is a combination of the organisation’s official abbreviation (UNESCO) and the second part of the genus Pontonia Latreille, 1829 , the nominal genus of the formerly recognised palaemonid subfamily Pontoniinae , which included all symbiotic palaemonid genera. Genus feminine.
Remarks. The combination of the following three characters, viz. (1) the presence of strong supraorbital teeth on the carapace; (2) the presence of a double-fossa system on the fingers of the major second pereiopod; and (3) the presence of a row of spinules (spiniform setae) at the lateral angle of the diaeresis of the uropodal exopod, separates Unesconia gen. nov. from all other “pontoniine” genera currently known from the eastern Pacific (Table 1). Only two other genera present in the eastern Pacific include species associated with sponges (Demospongiae), viz. Periclimenaeus Borradaile, 1915 and Typton Costa, 1844 . However, Unesconia gen. nov. differs from both of them by a number of morphological features, for instance, on the major second pereiopod (major cheliped). In Periclimenaeus , the major cheliped is characterised by the presence of a strong tooth (= plunger) on the dactylus and a deep fossa on the pollex, whereas in Typton , the fingers are devoid of fossae (e.g. Holthuis 1951; Chace 1972; Pachelle et al. 2015), both of these configurations contrasting with a double-fossa system on the fingers in the new genus. Unesconia gen. nov. also differs from Periclimenaeus and Typton by several additional features on the frontal margin of the carapace, first pereiopod, ambulatory pereiopods and uropodal exopod (e.g. Holthuis 1951; Chace 1972; Pachelle et al. 2015).
Unesconia gen. nov. also clearly differs from all western Atlantic palaemonid genera with sponge symbionts, including the monotypic Holthuisaeus Anker & De Grave, 2010 , presently containing only H. bermudensis ( Armstrong, 1940) , a relatively large shrimp characterised by the presence of a strong plunger on the pollex and a deep fossa on the dactylus (inverted snapping mechanism) ( Anker & De Grave 2010).
The phylogenetic placement of Unesconia gen. nov. within the Palaemonidae is not immediately clear. The total number of palaemonid genera containing both known or presumed sponge associates is 24 ( Bruce 2010; Marin & Chan 2013), including the herein described new genus, but excluding Isopericlimenaeus Marin, 2012 , recently synonymised with Periclimenaeus ( Park et al. 2019) . The vast majority of the sponge-associated palaemonid genera occur in the Indo-West Pacific, but most of them are morphologically very different from Unesconia gen. nov. Nevertheless, the new genus may be more closely related to a small Indo-West Pacific generic complex formed by Paraclimenaeus Bruce, 1988 , Apopontonia Bruce, 1976 and Climeniperaeus Bruce, 1996 ( Bruce 1976, 1988 a, 1988b, 1996, 2001, 2009; Mitsuhashi & Chan 2008; Odijk & Fransen 2017). The relationships between these three genera and the much larger and the morphologically heterogeneous Periclimenaeus (present also in the eastern Pacific and Atlantic) remain uncertain. In a recent phylogenetic analysis based on 16S gene sequences ( Odijk & Fransen 2017), Periclimenaeus was represented by only two terminals, whereas in the more comprehensive analysis of the symbiotic palaemonid shrimps based on four genes (COI, 16S, H 3, 18S) performed by Horká et al. (2016), Periclimenaeus was represented only by one terminal, with no representatives of Paraclimenaeus , Apopontonia and Climeniperaeus .
Paraclimenaeus , Apopontonia and Climeniperaeus are united by the well-developed, dorsally dentate rostrum, with expanded lateral carinae; the carapace without hepatic tooth; all maxillipeds with well-developed exopods; the fourth thoracic sternite without median process; the dactyli of the third to fifth pereiopods biunguiculate and with the corpus ventrally dentate; and the uropodal exopod with a row of spinules on the lateral margin ( Bruce 2009; Odijk & Fransen 2017). Although Unesconia gen. nov. generally fits in this generic group, it can be separated from these three genera by several morphological characters. In Unesconia gen. nov., the corpus of the ambulatory (third to fifth) pereiopod dactyli bears one large distal tooth and two or three much smaller, apparently mobile spinules along its ventral margin. This configuration of the ambulatory dactylus is also seen in Apopontonia , although the dactylus itself is much stouter (cf. Bruce 1976: fig. 5I). In contrast, in Paraclimenaeus and Climeniperaeus , the corpus of the ambulatory dactylus possesses five or more accessory teeth, including the larger distal tooth (cf. Bruce 1988a: fig. 6D; Bruce 1988b: fig. 7G; Odijk & Fransen 2017: fig. 5B, D, F).
In Unesconia gen. nov., the uropodal exopod typically bears five spinules at the lateral portion of the diaeresis, with the median or central spinule being conspicuously longer than the others; none of the spinules extends over the well-marked distolateral angle of the exopod. This configuration is similar to that of Apopontonia , which has seven spinules, one of them noticeably longer ( Bruce 1976: fig. 3F), although not as long as in the new genus. The uropodal exopod of Paraclimenaeus and Climeniperaeus is markedly different from that of Unesconia gen. nov. and Apopontonia : the spinules are much more numerous (around 10 or more), the longest is typically the most mesial one (or one of the mesial ones), in addition being only slightly longer than the others; most importantly, they extend over the poorly marked, rounded edge of the diaeresis onto the lateral margin of the exopod (cf. Holthuis 1952b: fig. 48c; Bruce 1988a: fig. 6K; Bruce 1988b: fig. 5G; Odijk & Fransen 2017: fig. 2E).
Paraclimenaeus differs from Unesconia gen. nov. by the absence of supraorbital teeth (cf. Bruce 1988a: fig. 2B; Odijk & Fransen 2017: fig. 2A), which are very prominent in the type species of the new genus. Apopontonia , now restricted to the type species, Apopontonia falcirostris Bruce, 1976 , and Climeniperaeus , currently with three species ( Bruce 1976, 2009; Odijk & Fransen 2017), both have prominent orbital teeth (cf. Holthuis 1952b: fig. 48b; Bruce 1976: fig. 3B; Bruce 1988b: fig. 5B; Bruce 1996: fig. 4). However, only Apopontonia has a well-developed rostral carina, in addition to the supraorbital teeth, thus approaching most the general shape of the fronto-orbital region of Unesconia gen. nov.
Most species of Paraclimenaeus and Climeniperaeus can also be separated from the new genus by the presence of a relatively stout tooth on the pollex and a deep fossa on the dactylus of the major second pereiopod, as in Holthuisaeus (inverted snapping mechanism, cf. Bruce 1996: fig. 4E), although in some species currently assigned to Paraclimenaeus , the tooth of the pollex appears to be much less developed and excavated, and is thus reminiscent of the two opposing fossae seen in the new genus (cf. Odijk & Fransen 2017: p. 1109, although not clear in their fig. 4C).
Taking all characters into account, Unesconia gen. nov. appears to be morphologically closest to Apopontonia , especially in the shape of the rostro-orbital area of the carapace, ambulatory dactylus and uropodal exopod. The general shape of the telson, including the armature of the dorsal surface and posterior margin, are also remarkably similar in the two genera (cf. Bruce 1976: fig. 3C). However, Apopontonia differs from the new genus by the slightly unequal and subsymmetrical second pereiopods, devoid of fossae or any other snapping mechanism on the chela fingers (cf. Bruce 1976: figs. 2, 5F). Initially, the present author considered the possibility that Bruce’s (1976) single holotype specimen of A. falcirostris , an ovigerous female, had abnormal second pereiopods, e.g. one of them (major) was regenerating. However, this possibility can be excluded since subsequently other specimens of the species that agreed very closely with the original description have been reported ( Bruce 1981; Marin 2007), i.e. they must have had subsymmetrical second pereiopods, as described and illustrated by Bruce (1976). Another important character that separates Unesconia gen. nov. from Apopontonia , Paraclimenaeus and Climeniperaeus is the first pereiopod chela with broadened and deeply excavated fingers, with both dactylus and pollex ending in three sharp teeth ( Fig. 4D, E View FIGURE 4 ). This feature is absent or at least has not been illustrated in the very detailed taxonomic accounts of Holthuis (1952b), Bruce (1976, 1988 a, 1988b, 2009) and Odijk & Fransen (2017). For instance, the first pereiopod fingers of A. falcirostris are neither broadened nor excavated, and their tips are simple, not tridentate (cf. Bruce 1976: fig. 5C).
Without a comprehensive phylogenetic analysis, it would be premature to further speculate on the phylogenetic position of Unesconia gen. nov. and also on how the symbiosis with sponges may have evolved in this new palaemonid lineage. Host switches occur widely within the Palaemonidae , as exemplified by the switches between the tunicate and bivalve mollusc hosts in the clade containing Pontonia Latreille, 1829 , Ascidonia Fransen, 2002 and several allied Indo-West Pacific genera ( Fransen 2002). Marin & Anker (2008) hypothesised a further host switch from bivalve molluscs to tunicates within the genus Pontonia . Horká et al. (2016) found multiple events of host switching in the Palaemonidae , including to and from demosponges, but also retrieved a small clade containing predominantly sponge-associated taxa, e.g. Periclimenaeus and several other exclusively sponge-associated genera (part of their clade 5 in fig. 4). Within this clade, at least one host switch occurred from sponges to tunicates within Periclimenaeus , which is possibly the morphologically closest genus to Unesconia gen. nov. in the eastern Pacific. Whether Unesconia gen. nov. is indeed related to Periclimenaeus (even more distantly) or whether it belongs to the clade 5 of Horká et al. (2016) together with other sponge associates currently remains unknown. Future phylogenetic analyses should include representatives of Unesconia gen. nov., Paraclimenaeus , Apopontonia , Climeniperaeus , as well as a much wider selection of taxa currently assigned to Periclimenaeus , Typtonomenaeus Marin & Chan, 2013 and Anisomenaeus Bruce, 2010 , the latter two with clear affinities to Periclimenaeus , to elucidate the evolution of this interesting shrimp lineage.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.