
Magelona pacifica Monro, 1933
|
publication ID |
https://doi.org/ 10.5281/zenodo.208658 |
|
DOI |
https://doi.org/10.5281/zenodo.5658293 |
|
persistent identifier |
https://treatment.plazi.org/id/EA76A055-FFBD-FFA6-FF0A-D3610B3EFA05 |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona pacifica Monro, 1933 |
| status |
|
Magelona pacifica Monro, 1933 View in CoL
Figures 14–16 View FIGURE 14 View FIGURE 15 View FIGURE 16
Magelona pacifica Monro, 1933: 1048 View in CoL –1049, fig. 2 Material examined. Syntypes: PANAMÁ, Gorgona Island, at low tide, collected by Dr. C. Crossland, (BMNH Type 1933.7.10.65/70; 14 af, 13 f of 2–32 chaetigers, 1 pf of approximately 28 chaetigers for 9 mm, and 13 loose palps) (N.B. original description indicates 12 specimens).
Diagnosis. Prostomium longer than wide, with distinct prostomial horns. Notopodia of chaetigers 1–8 with spatulate postchaetal lamellae, expanded as cirriform dorsal superior processes. Neuropodia with slender triangular ventral lobes; chaetiger 8 with additional triangular postchaetal lamellae. Notopodia of chaetiger 9 with rounded postchaetal lamellae confluent with prechaetal ridges, expanded as lateral cirriform lobes. Neuropodia of chaetiger 9 with triangular postchaetal lamellae and ventral prechaetal processes. All thoracic chaetae capillary. Abdominal lateral lamellae rounded triangular (toad-stool shape in profile), basally constricted. Hooded hooks bidentate, in one unidirectional group.
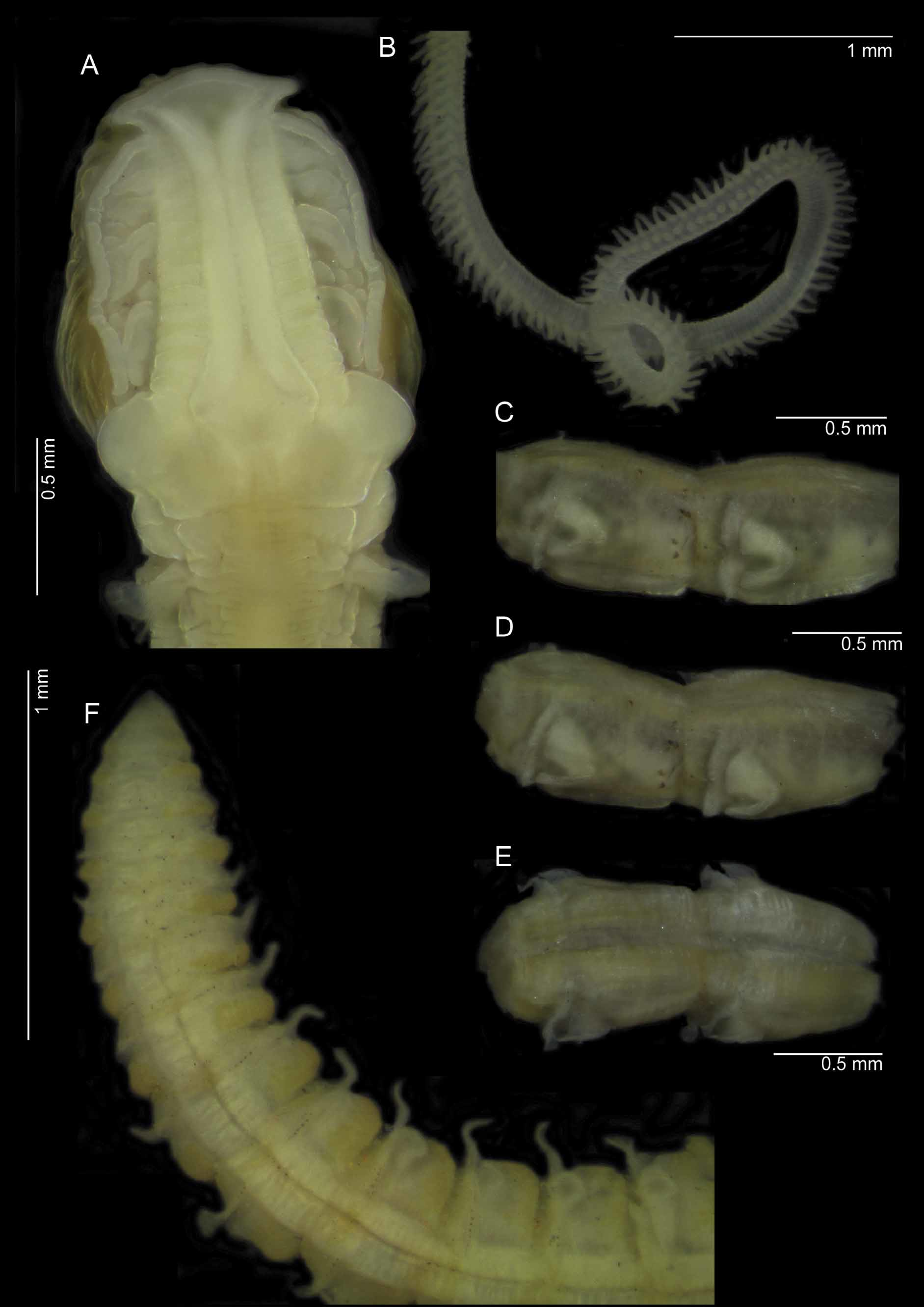
Re-description. A moderately large species ( Figures 14 View FIGURE 14 A–D); junction between thorax and abdomen distinct, abdomen thicker dorso-ventrally than thorax. All specimens posteriorly incomplete, condition generally poor. Thoracic parapodia missing or incomplete in many specimens, particularly in posterior thorax. Dimensions of longest syntype: prostomium 1.0 mm long, 0.8 mm wide; thorax 5.0 mm long (including prostomium), 0.5 mm wide (thorax generally widest at chaetiger 1, tapering towards abdomen); abdomen 0.4 mm wide; total length approximately 33.5 mm for 60 chaetigers. Other syntypes with 29–51 chaetigers for 15–26 mm (N.B. original description states that “one of the most complete specimens measures 45 mm (…) for 85 chaetigers”. It is likely that this specimen has now fragmented and may be represented by the 60 chaetiger af and the posterior fragment reported here).
Prostomium longer than wide (L:W ratio 1.1–1.4); anterior margin smooth, triangular, with very conspicuous prostomial horns ( Figure 15 View FIGURE 15 A) (the tips of which are more rounded, and the horns broader than originally figured by Monro). Prostomial lateral margins often undulating. Two pairs of prominent longitudinal dorsal muscular(?) ridges, outer pair abutting inners for entire length; inner pair diverging distally into each horn. Outer pair moderately thick, heavily ridged transversely. Inner pair showing diagonal striations at each end, with light transverse ridging medially. Distinct muscular(?) areas either side of ridges of varying shape and size. Proboscis everted in 13 specimens; heart-shaped when fully everted, round to oval ( Figure 14 View FIGURE 14 D) when partially everted; ridged longitudinally, although much lighter superiorly, appearing almost smooth. Palps attached on eight specimens, long and slender; arising ventrolaterally from base of prostomium, reaching approximately chaetiger 21–30; non-papillated region long reaching chaetiger 3–5. Degree of papillation variable, papillae short proximally but long for majority of length; digitiform; proximally with two or three rows of papillae either side of inconspicuous groove, medially with two, and distally with one or two rows either side.
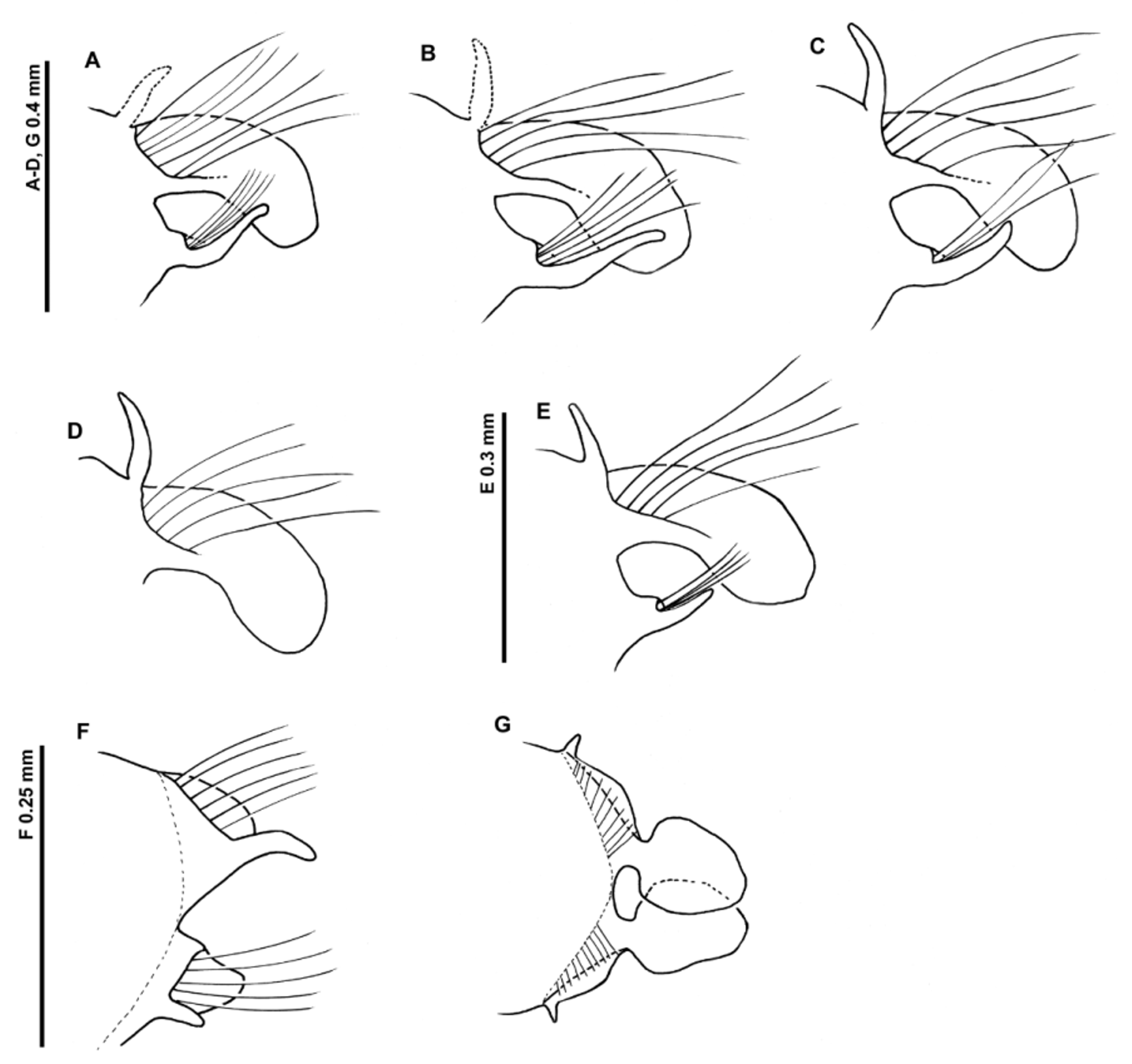
Achaetous region behind prostomium large approximately one and a half times the size of chaetiger 1. Chaetigers 1–7 similar (although size of the lamellae and processes in posterior thorax difficult to ascertain due to absence/damage in most specimens); parapodia biramous ( Figures 16 View FIGURE 16 A–E); notopodia with low triangular prechaetal lamellae confluent with large spatulate postchaetal lamellae, of a similar size throughout the thorax, upper margins smooth. Long slender, cirriform, prechaetal superior processes present (DML). Neuropodial pre- and postchaetal lamellae as low ridges, forming distinct cuff-like structures confluent with ventral triangular lobes (VNL) beneath chaetae, decreasing in size along thorax. Lamellae of chaetiger 8 unknown due to damage in all specimens, however, neuropodia with additional triangular postchaetal lamellae of similar size to those of the proceeding chaetiger.
Chaetiger 9: Notopodial prechaetal lamellae as low ridges confluent with rounded postchaetal lamellae (smaller than those of preceding chaetigers) and inferiorly expanded as small cirriform processes; superior processes (DML) not observed and described originally as absent ( Figure 16 View FIGURE 16 F). Neuropodia with triangular postchaetal lamellae confluent with low prechaetal ridges and digitiform prechaetal lobes. Chaetae of all thoracic chaetigers simple winged capillaries.
Abdominal chaetigers with large rounded triangular to spatulate lateral lamellae (toad-stool shape in profile) ( Figure 16 View FIGURE 16 G) (shape variable, sometimes more rounded, edges often undulating and folded over) of about equal size in both rami; basally constricted, stalked, and overlapping in anterior abdomen. Postchaetal extension of the lateral lamellae behind chaetal rows present in anterior abdomen; triangular, much reduced by chaetigers 15–20. Triangular processes (DML and VML) present at inner margins of chaetal rows, long in anterior abdomen.
Abdominal chaetae bidentate hooded hooks, with around 10–12 hooks per rami initially (not drawn to prevent further damage to specimens, particularly any remaining complete hooks). Majority of hooks broken where they emerge in all specimens, however they appear to be in one unidirectional group (difficult to discern due to condition of material. Monro’s original description does not mention this character, and although his drawings show unidirectional hooks, they are shown with different orientations in different rami i.e. facing the lamellae in one group and away in another). A parapodial slide preparation made from a posterior - most chaetiger of a syntype specimen suggests that they are unidirectional based on how they arise. Information on non - type material of M. pacifica from Mexico also suggests unidirectional facing bidentate hooks ( Brasil 2003).
Paired posteriorly open pouches on consecutive segments, on both sides of the body, starting from around chaetigers 36–40 (observed on 10 specimens) are present. Pouches appear as simple folds, medially split with thicker cuticle surrounding edges ( Figures 15 View FIGURE 15 C–E). Pouches present until approximately 10 chaetigers from the pygidium, (observed from posterior fragment).
Eggs present in three fragments (18 and 20 chaetiger fragments and 44 chaetiger af), approximately 80 Μm in diameter (clearly visible from approximately 37th chaetiger). Posterior fragment ( Figure 15 View FIGURE 15 F) with no pygidial cirri observed, presumed broken(?).
Colour. Colour of preserved specimens uniformly cream in alcohol. Darker colouration interparapodially in the abdomen ( Figure 15 View FIGURE 15 F). Methyl green stain dissipates quickly leaving a very diffuse, overall stain ( Figures 14 View FIGURE 14 C– D) (stain slightly darker and persisting longer on chaetigers 1–7 dorsally and 3–7 ventrally). However, dorsal transverse bands of light green/yellow persist between chaetigers 2–9, just posterior to parapodia. Those behind chaetiger 2 often appearing as patches rather than bands. These bands correspond with light speckled transverse bands visible in the same area without staining. An additional medial dorsal patch, just prior to chaetiger 1 is often present. Similar light green bands observed ventrally between 4–9.
Habitat. Type specimens found at low tide on Gorgona Island, Panamá.
Distribution. Magelona pacifica has been additionally recorded from Southern California ( Hartman 1944; 1956; 1969), Gulf of California and the Pacific coast of México ( Arias-González 1984; Padilla-Galicia 1984; Varela-Hernández 1993; De León-González 1994; Hernández-Alcántara & Solís-Weiss 2000). Ten further records were made throughout the Tropical Eastern Pacific (see, Salazar-Vallejo & Londoño-Mesa 2004), and also West of the Philippine Trench (10°26'N, 126°05'E, 1000 m) ( Kirkegaard 1996). Kirkegaard’s description states “without notopodial and neuropodial medial lamellae in posterior region” as in the original description by Monro (1933): “I can see no cirriform processes”. However, Magelona pacifica has been shown herein to possess abdominal triangular processes (DML and VML) at the inner margins of chaetal rows, so we cannot be sure whether Kirkegaard’s material is actually M. pacifica . Paxton & Chou (2000) report the presence of Magelona cf. pacifica from the South China Sea.
Remarks. The poor condition of the material examined herein makes a full re-description of this species from type material difficult, particularly the parapodia between chaetigers 5–8. Examination of newly collected material from the type locality would be prudent. The species sharing the most morphological similarities with M. pacifica is M. montera sp. nov., and their affinities are discussed above for the justification of the new species. Several features differ between Monro’s original description and this re-description. In particular, the presence of triangular processes at the inner margins of chaetal rows (DML and VML) in the abdomen. Also, the presence of triangular postchaetal lamellae in the neuropodia of the 8th chaetiger were not mentioned in the original description.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Magelona pacifica Monro, 1933
| Mortimer, Kate, Cassà, Susanna, Martin, Daniel & Gil, João 2012 |
Magelona pacifica
| Monro 1933: 1048 |