
Cobbionema acuminata, Ahmed & Boström & Holovachov, 2020
|
publication ID |
https://doi.org/ 10.5852/ejt.2020.702 |
|
publication LSID |
lsid:zoobank.org:pub:B4DDC9C7-69F4-40D1-A424-27D04331D1F8 |
|
DOI |
https://doi.org/10.5281/zenodo.4328585 |
|
persistent identifier |
https://treatment.plazi.org/id/AF9C8B50-B488-4425-A883-8A1ACAB423E7 |
|
taxon LSID |
lsid:zoobank.org:act:AF9C8B50-B488-4425-A883-8A1ACAB423E7 |
|
treatment provided by |
Valdenar |
|
scientific name |
Cobbionema acuminata |
| status |
sp. nov. |
Cobbionema acuminata sp. nov.
urn:lsid:zoobank.org:act:AF9C8B50-B488-4425-A883-8A1ACAB423E7
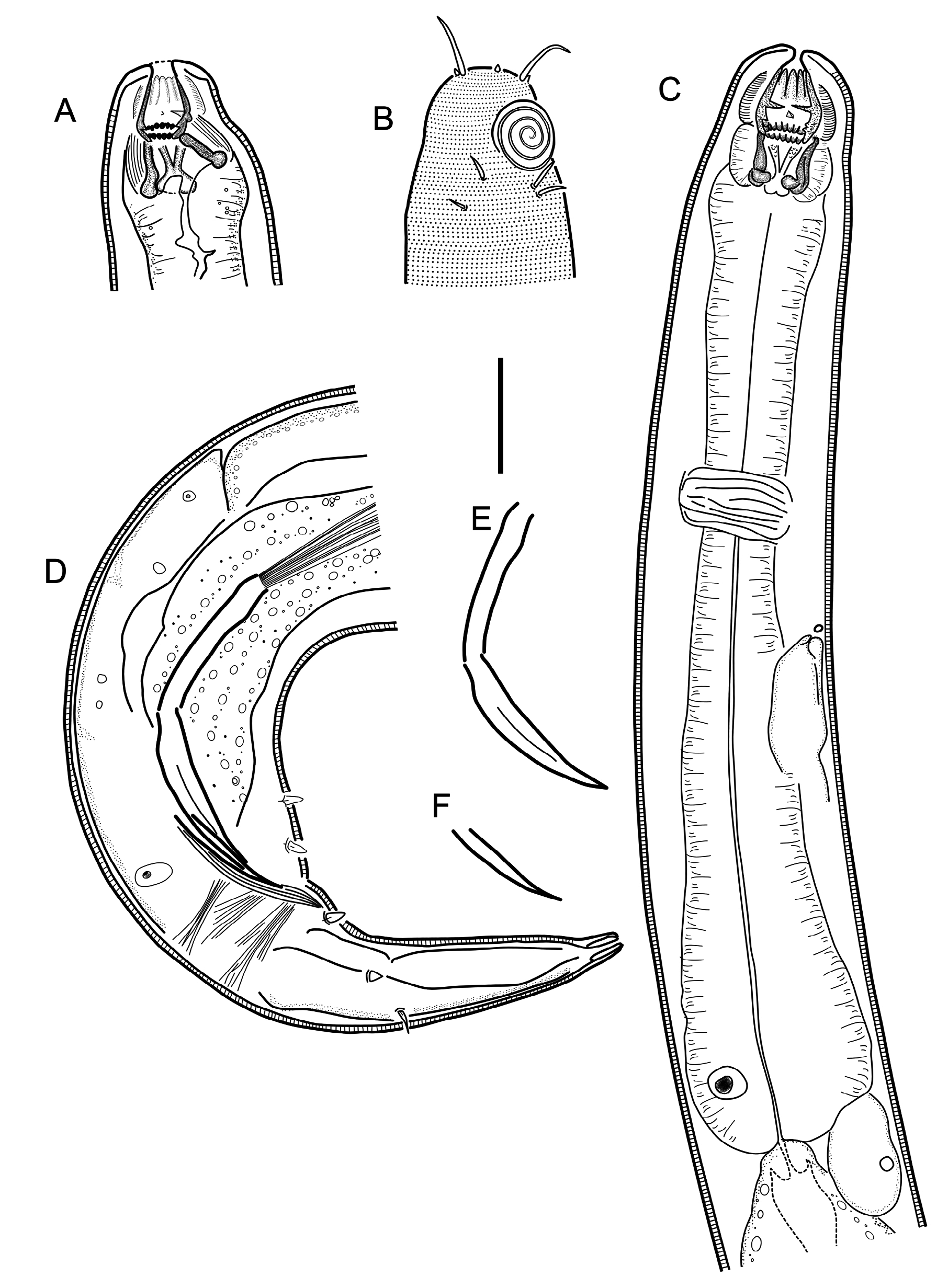
Figs 8–9 View Fig View Fig , Table 2
Diagnosis
Cobbionema acuminata sp. nov. is characterised by 1.09–1.37 mm long body; lateral alae and body pores absent; cephalic sensilla equal to 0.6–0.8 labial region diameters in length; amphideal fovea large, with 3.75 turns; two rows of pointed projections and three prominent sharply pointed tines in the stoma, rows of projections wider apart longitudinally; spicules consisting of two sections, 55–65 µm long; single midventral precloacal papilliform sensillum; tail curved ventrally, conical in shape.
Etymology
The species name refers to the presence of acute, sharply pointed tines in the stoma.
Material examined
Holotype
SWEDEN • ♂; Skagerrak , Hållö island; 58º20.35′ N, 11º12.70′ E; 19 Jul. 2014; 14–17 m depth; O. Holovachov leg.; sand and shells; SMNH Type-9221 . GoogleMaps
Paratype
SWEDEN • 1 ♂; same collection data as for holotype; SMNH Type-9222 GoogleMaps .
Additional non-type material
SWEDEN • 1 ♂; Skagerrak , Bonden island; 58º12.64′ N, 11º18.95′ E; 10 Nov. 2012; 15–22 m depth; O. Holovachov leg.; sand and shells; SMNH-182043 GoogleMaps .
Description
Adult characters
Body cylindrical, tapering slightly towards both extremities. Cuticle with transverse rows of dots, without lateral differentiation. Lateral alae absent. Body pores absent. Somatic setae scattered along the body. Labial region bluntly-rounded, not set-off, but cephalic region demarcated by a broad and shallow constriction at level with amphidial fovea. Six equal lips surrounding mouth opening. Inner labial sensilla indistinct. Outer labial sensilla small papilliform, located on the periphery of labial region. Cephalic sensilla setiform, equal to 0.6–0.8 labial region diameters in length, located at the same level as outer labial sensilla. Amphideal fovea ventrosublateral, large, multispiral with 3.75 turns, with circular outline, equal to about 0.6 of the corresponding body diameter. Four pairs of cervical setae present, located just posterior to amphid in ventrosublateral (at the base of each amphid) and dorsosublateral positions ( Fig. 8B View Fig ). Buccal cavity voluminous, divided into anterior and posterior chambers. Anterior chamber of buccal cavity barrel-shaped, with 12 cuticularized rhabdions, each with two pairs of large, pointed projections at posterior extremity. These projections are arranged in two horizontal rows, appearing from an en face view as two circles of denticles separating stoma into anterior and posterior chamber. Three (one middorsal and two ventrosublateral) prominent and sharply pointed tines (with hollow conoid base) located a short distance in front of anterior circle of denticles. Posterior buccal chamber cone-shaped, widest at (posterior or anterior) extremity (depending on the state of stomatal musculature), with strongly cuticularized Y-shaped mandibles; anterior branches of each mandible with a field of minute denticles/knobs on its inner surface; posterior ‘stem’ of each mandible with strongly developed basal (submedian) knobs. Anterior buccal chamber surrounded by strongly developed sphincter muscle. Each mandible is also supported by a strong longitudinal and tangential musculature, altogether appearing as a muscular bulb. Pharynx weakly muscularised, with strongly developed glandular tissue throughout its entire length; with conspicuous anterior swelling surrounding the base of buccal cavity; pharynx widens posteriorly but not forming true posterior bulb. Cardia small, with cuticularized lumen. Secretory-excretory system present, renette body located at level with pharyngointestinal junction, secretory-excretory pore located short distance posterior to nerve ring. Tail curved ventrally, conical in shape. Caudal glands present, opening via a common spinneret, caudal gland cells/ bodies at level with posterior part of intestine, anterior to cloaca.
Male
Reproductive system diorchic, with outstretched anterior testis and reflexed posterior testis. Anterior testis on right side of intestine, posterior testis on left side of intestine. Spicules paired and symmetrical, elongate, seemingly consisting of two sections separated by a constriction, subcylindrical/fusiform proximal section and conoid distal section, equal to 1.9–2.4 anal body diameters in length. Gubernaculum reduced only to sublateral crura, arcuate conoid in shape, flanking distal part of spicules. Single midventral precloacal papilliform sensillum located 17 µm in front of cloacal opening ( Fig. 8D View Fig ). Further six papilliform sensilla arranged as follows: one subventral pair located half-way between midventral papilliform sensillum and cloacal opening; one subventral pair short distance posterior to cloalcal opening and one ventro-sublateral pair at anterior third of tail length. One pair of dorsosublateral setae located along middle third of tail.
Female
Not found.
Differential diagnosis
Cobbionema acuminata sp. nov., known from three males, differs from all previously described species in arrangement of the stoma armaments (rows of pointed projections and tines widely spaced longitudinally vs rows of projections and tines closer longitudinally in all other species, the three sharply pointed tines more erect and prominent vs pointed posteriorly and shorter in other species). Cobbionema acuminata sp. nov. is also unique in its lack of body pores (present in all other species except for C. capense where no mention was made of the presence or absence of body pores in the description). Cobbionema acuminata sp. nov. differs from males of C. acrocerca and C. cylindrolaimoides in its tail shape (conical in C. acuminata sp. nov. vs conoid proximal and digitate distal sections in C. acrocerca and C. cylindrolaimoides ) and lacking lateral alae (present in C. acrocerca and C. cylindrolaimoides ). Cobbionema acuminata sp. nov. also differs from C. acrocerca in most body measurements (see Table 2). Cobbionema acuminata sp. nov. differs from holotype and paratype males of C. trigamma in its body diameter (25.9–31.5 vs 16 and 17 in holotype and paratype of C. trigamma ), number of pointed projections in the stoma (48 vs 24 in C. trigamma ) and tail shape (conical vs conoid proximal and digitate distal sections in C. trigamma ). Because C. capense is known only from a single female specimen, its comparison with C. acuminata sp. nov., for which no female specimens were found, cannot be based on a character with known sexual dimorphisms such as body size. The two can be distinguished based on their tail shape (conical vs conoid proximal and subcylindrical distal sections in C. capense ). Cobbionema acuminata sp. nov. resembles males of C. brevispicula sp. nov. in body length (1094–1366 µm vs 1083–1173 µm). The two, however, can be distinguished by their spicule shape and length (two-sectioned spicules consisting of subcylindrical proximal part and a conoid/fusiform distal section and measuring 1.9–2.4 anal body diameters long vs nearly straight, with bifid tips and measuring 0.9–1.0 anal body diameters long).
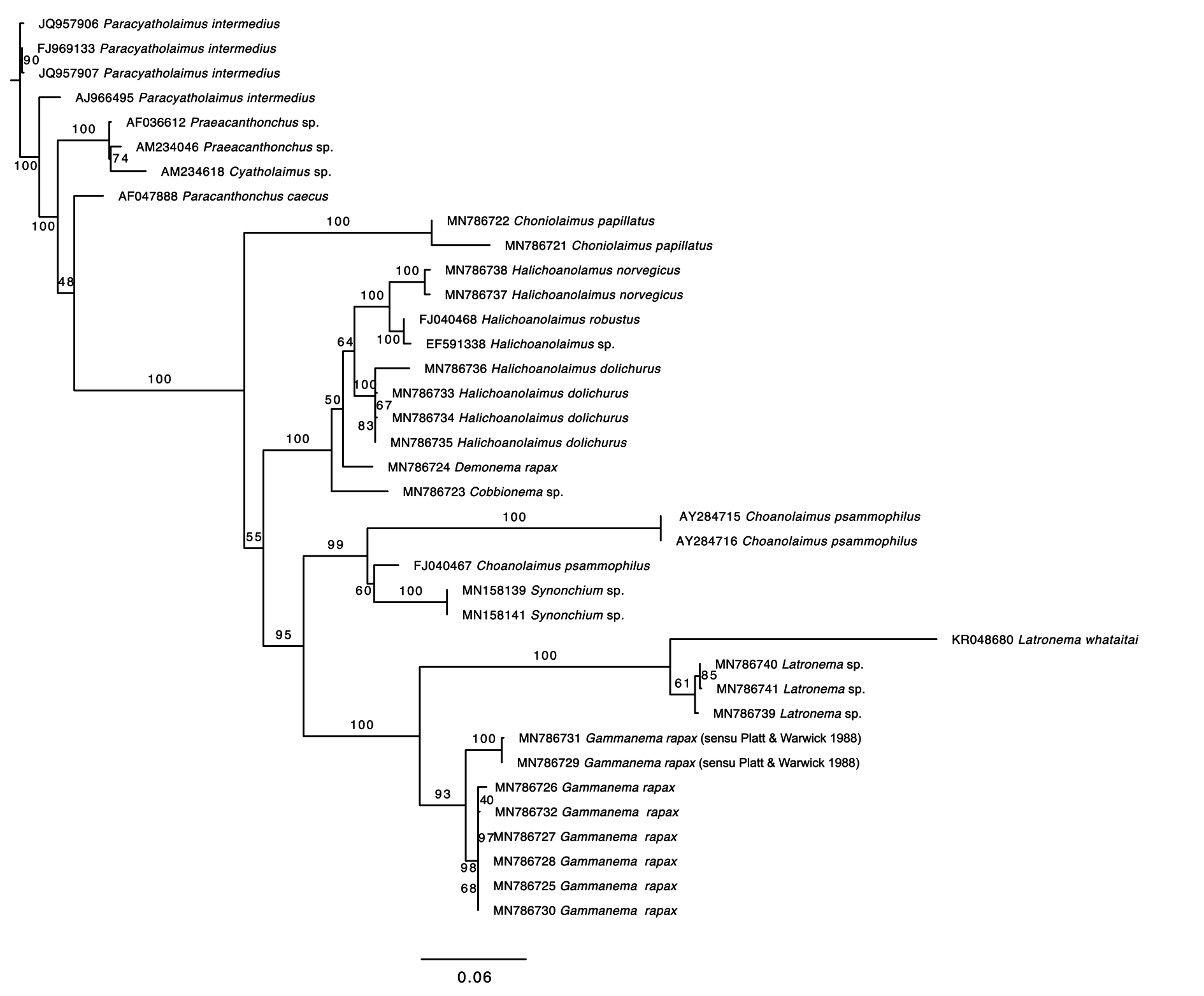
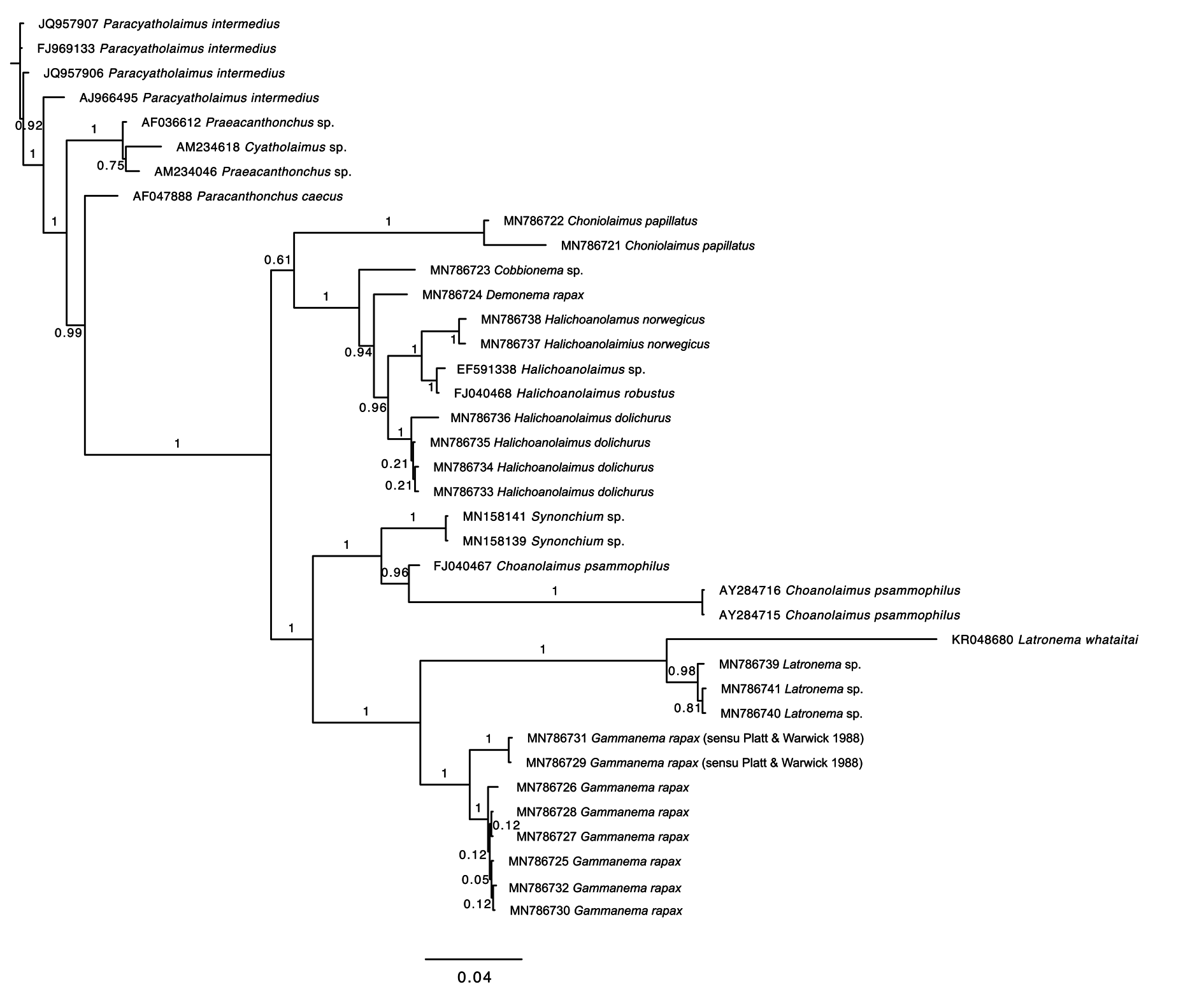
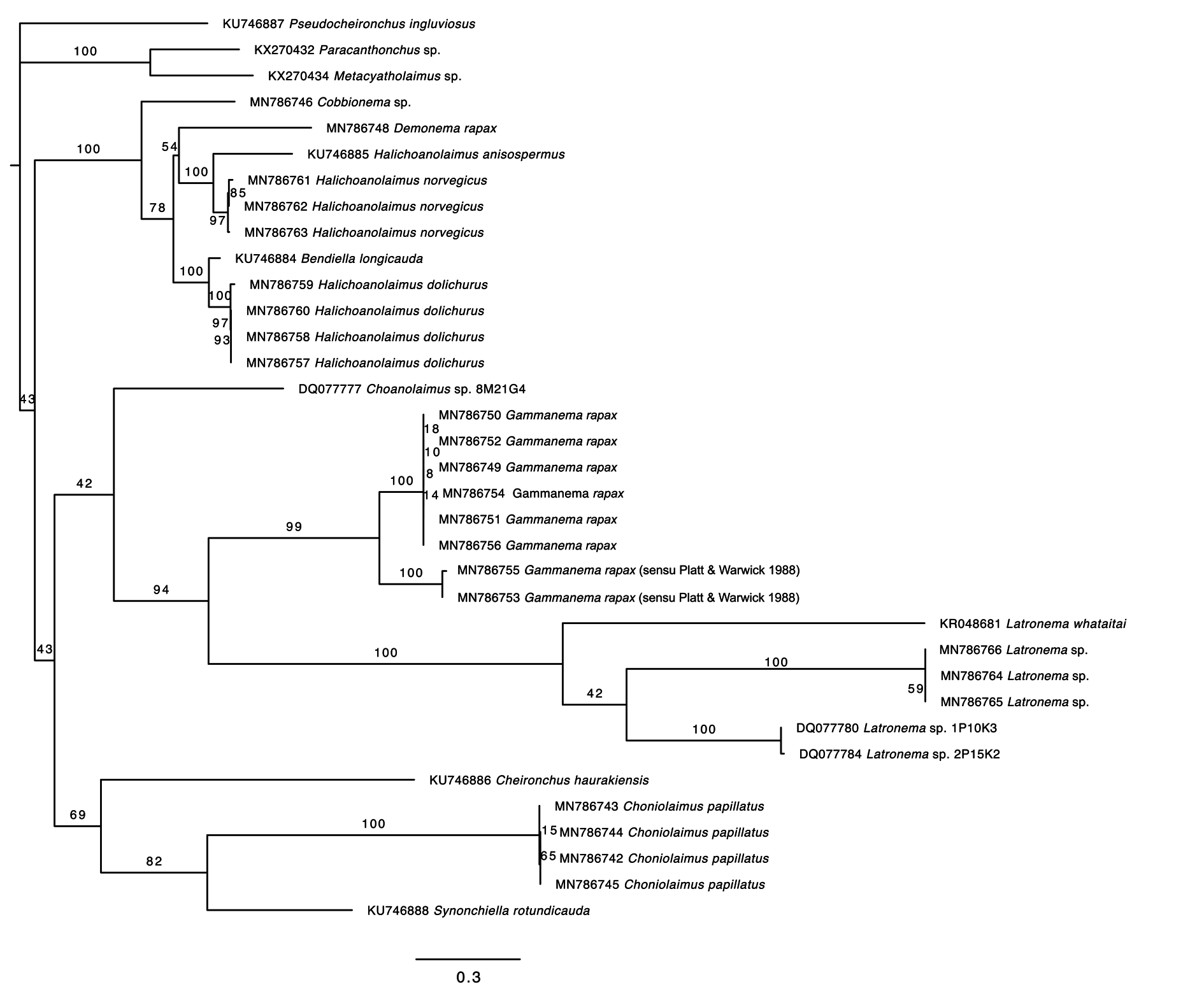
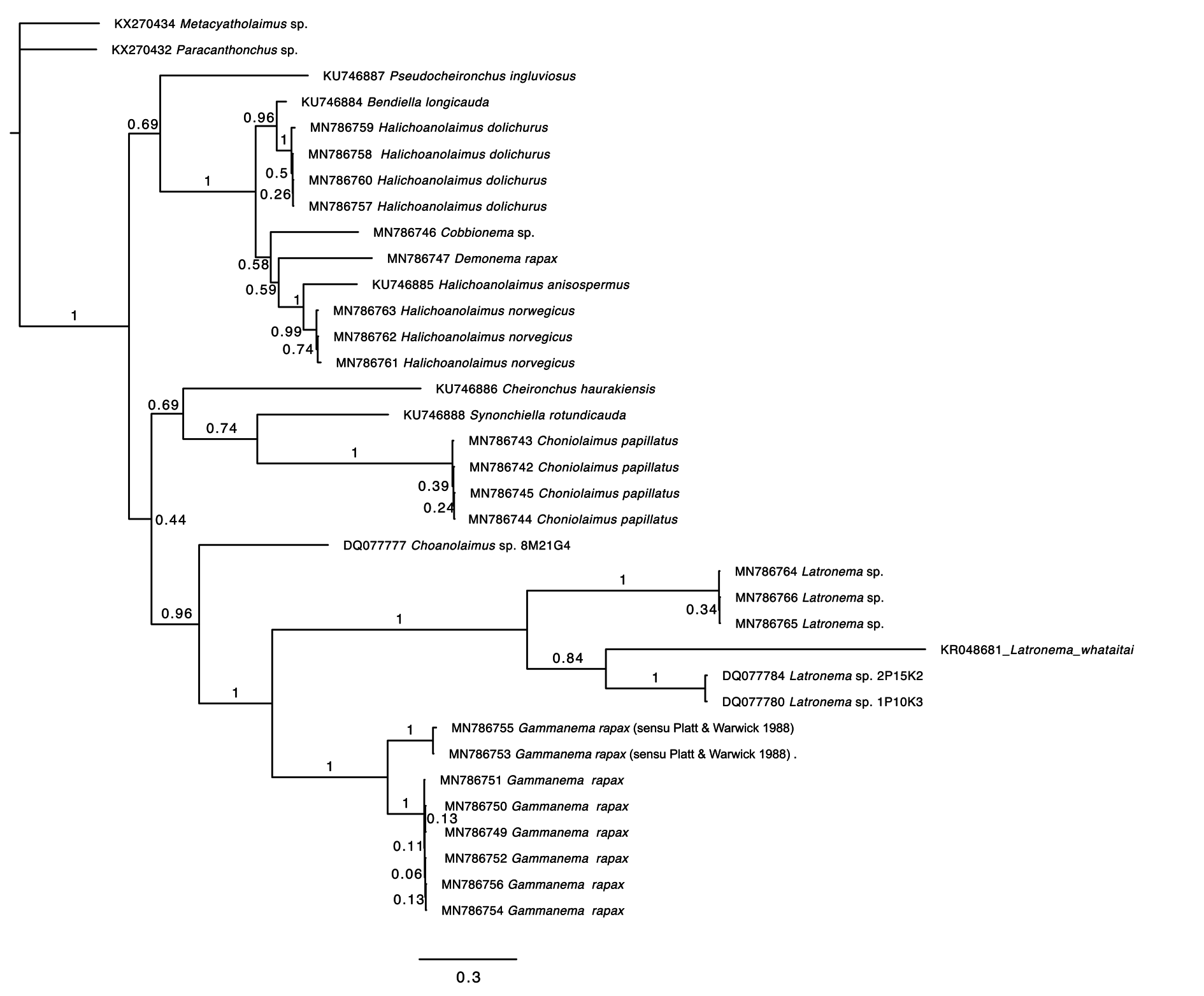
Molecular phylogeny
Both the 18S and 28S rRNA genes datasets were analysed using Maximum Likelihood (RAxML) and Bayesian inference methods (mcmcphase in PHASE 3). Analysis of the 18S rRNA dataset produced similar, but not identical tree topologies for the Maximum Likelihood and Bayesian inference methods (see Figs 10–11 View Fig View Fig ). The main topological differences between the two approaches concerned the relationship between Choniolaimus papillatus and the (( Halichoanolaimus + Demonema Cobb, 1894 ) + Cobbionema ) clade and the relationship between Choanolaimus psammophilus de Man, 1880 and Synonchium sp. Similarly, the 28S rRNA dataset produced tree topologies that were broadly comparable for the two inference methods (see Figs 12–13 View Fig View Fig ). The main difference between the tree topologies involves the position of Pseudocheironchus ingluviosus Leduc, 2013 , the position of Cobbionema within the ( Bendiella + Cobbionema + Demonema + Halichoanolaimus ) clade and the relationships within the Latronema Wieser, 1954 clade. All four trees indicate a close relationship between Latronema and Gammanema Cobb, 1920 through strong branch support (bootstrap support: 94–100 and posterior probability: 1). Cobbionema and Halichoanolaimus were nested within the same clade as was expected based on their morphological similarities. Demonema was also embedded within the same clade.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |