
Caenoplana variegata ( Fletcher & Hamilton, 1888 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4731.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:BC349407-17A3-4FAF-AB01-38772AD5BF51 |
|
DOI |
https://doi.org/10.5281/zenodo.3648550 |
|
persistent identifier |
https://treatment.plazi.org/id/EF7787FF-FFF8-7B27-BACC-AC903FE1F86D |
|
treatment provided by |
Plazi |
|
scientific name |
Caenoplana variegata ( Fletcher & Hamilton, 1888 ) |
| status |
|
Caenoplana variegata ( Fletcher & Hamilton, 1888)
Geoplana variegata Fletcher & Hamilton, 1888
Geoplana variegata Dendy, 1892
Geoplana variegata Dendy, 1893a
Geoplana variegata Dendy, 1893b
Geoplana variegata Dendy, 1894
Geoplana variegata Graff, 1899
Geoplana variegata Steel, 1900
Geoplana varigata Winsor, 1973
Geoplana varigata Winsor, 1977
Australopacifica bicolor Ogren & Kawakatsu, 1991
Australopacifica variegata Ogren & Kawakatsu, 1991
Caenoplana bicolor Winsor, 1991
Caenoplana variegata Winsor, 1991
Caenoplana bicolor Winsor, 1997
Caenoplana bicolor Álvarez-Presas et al. 2014
Espèce “rayée jaune” Justine et al., 2014
Caenoplana bicolor De Waart, 2016
Caenoplana bicolor Sluys, 2016
Caenoplana bicolor Vardinoyannis & Alexandrakis, 2019
Material examined. NHMUK.97.11.1.26-28, as Geoplana variegata, Bundanoon , NSW, T. Steel, 1897 [34.66S, 150.28E]. The selected specimen (henceforth referred to as NHMUK.97) had a prominent gonopore and faint visible dark lines dorsally ( Fig. 5a, b View FIGURE 5 ). It was cut into six portions: anterior portion 42 mm long and 30 mm posterior portion, neither sectioned (subsequently the posterior 5 mm severed and used for attempted molecular analysis); 4 mm pre-pharyngeal portion, TS, 7 slides; 13 mm long pharyngeal portion GoogleMaps , LS, 30 slides; 10 mm portion, not sectioned; 11 mm portion including the copulatory apparatus, LS, 27 slides.
Zoological Museum, Berlin ( ZMB) Vermes 3506; Geoplana bicolor , syntype (hereafter referred to as “ ZMB 3506”). This is one of the specimens found by W.B. Spencer in Queensland (Gympie; Burnett River; Cooran), Australia, mentioned in Dendy (1892) and sent by Dendy to Graff (Winsor, pers. comm). It has been sectioned by L. Winsor. 143 slides: Anterior, LS, 48 slides, H&E, 8μm; Mid-portion LS and TS, 47 slides, H&E (except 2 slides trichrome); genital portion, LS, 48 slides, H&E (except one trichrome).
NHMUK.2018.8.31.1. Collected near Coventry, UK [52.385449N, 1.648287W], 28 January 2015, J. Tomnie, (slides labeled and hereafter referred to as “Cv7”). It had a prominent gonopore thus mature. Killed in warm water for 30 s, then in 100% ethanol. Cut into five portions: the anterior 12 mm and posterior 8 mm ( Fig. 5 View FIGURE 5 , c & d) not sectioned and remain preserved in ethanol (the posterior tip was subsequently removed for molecular analysis); a 4 mm pre-pharyngeal portion, TS, 7 slides; an 8 mm pharyngeal section GoogleMaps , LS, 13 slides; a 12 mm portion containing the copulatory apparatus, LS, 16 slides.
Anatomical descriptions. External dimensions and the relative positions of the pharyngeal aperture (mouth) and gonopore of the three sectioned specimens, other available specimens and also taken from the literature are shown in Table 1 View TABLE 1 .
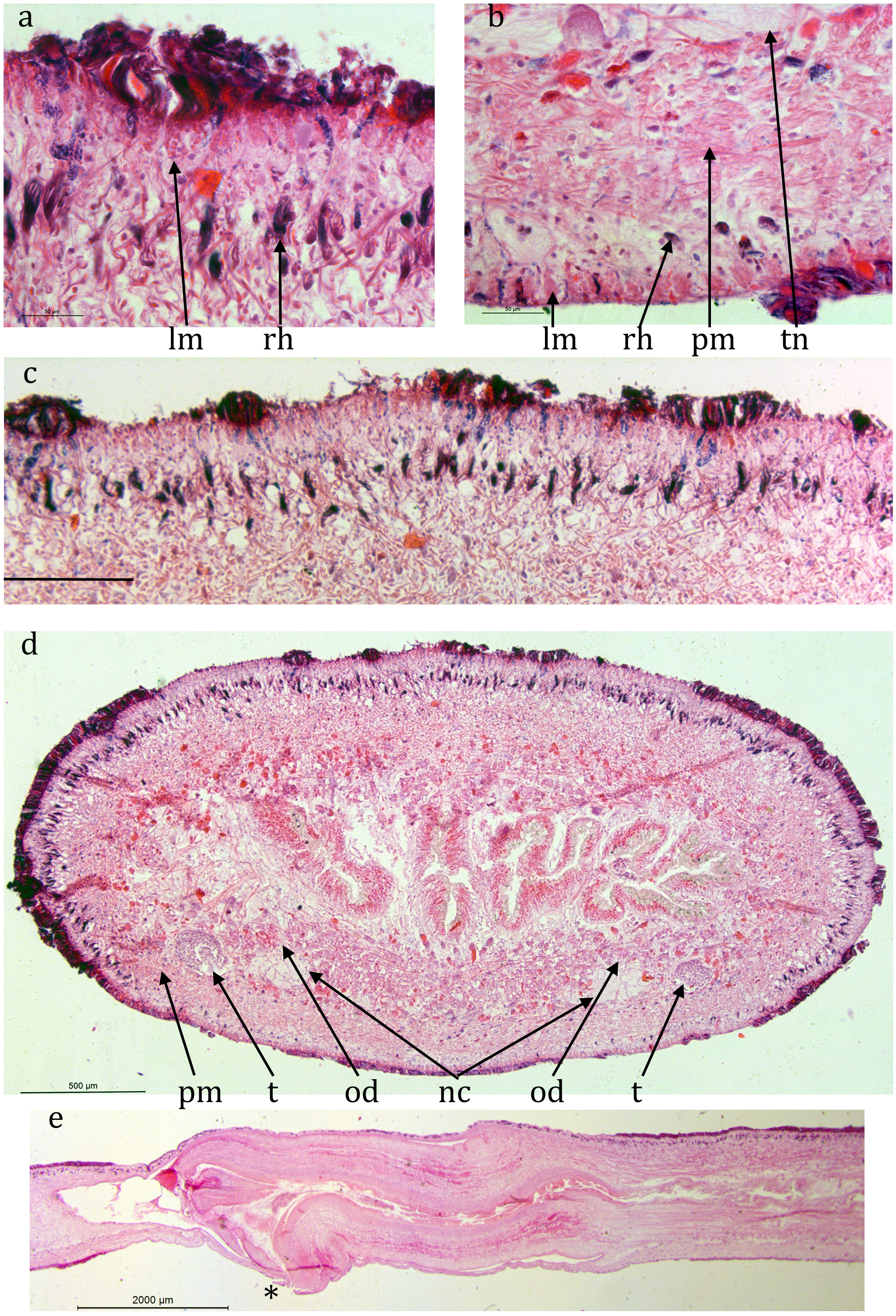
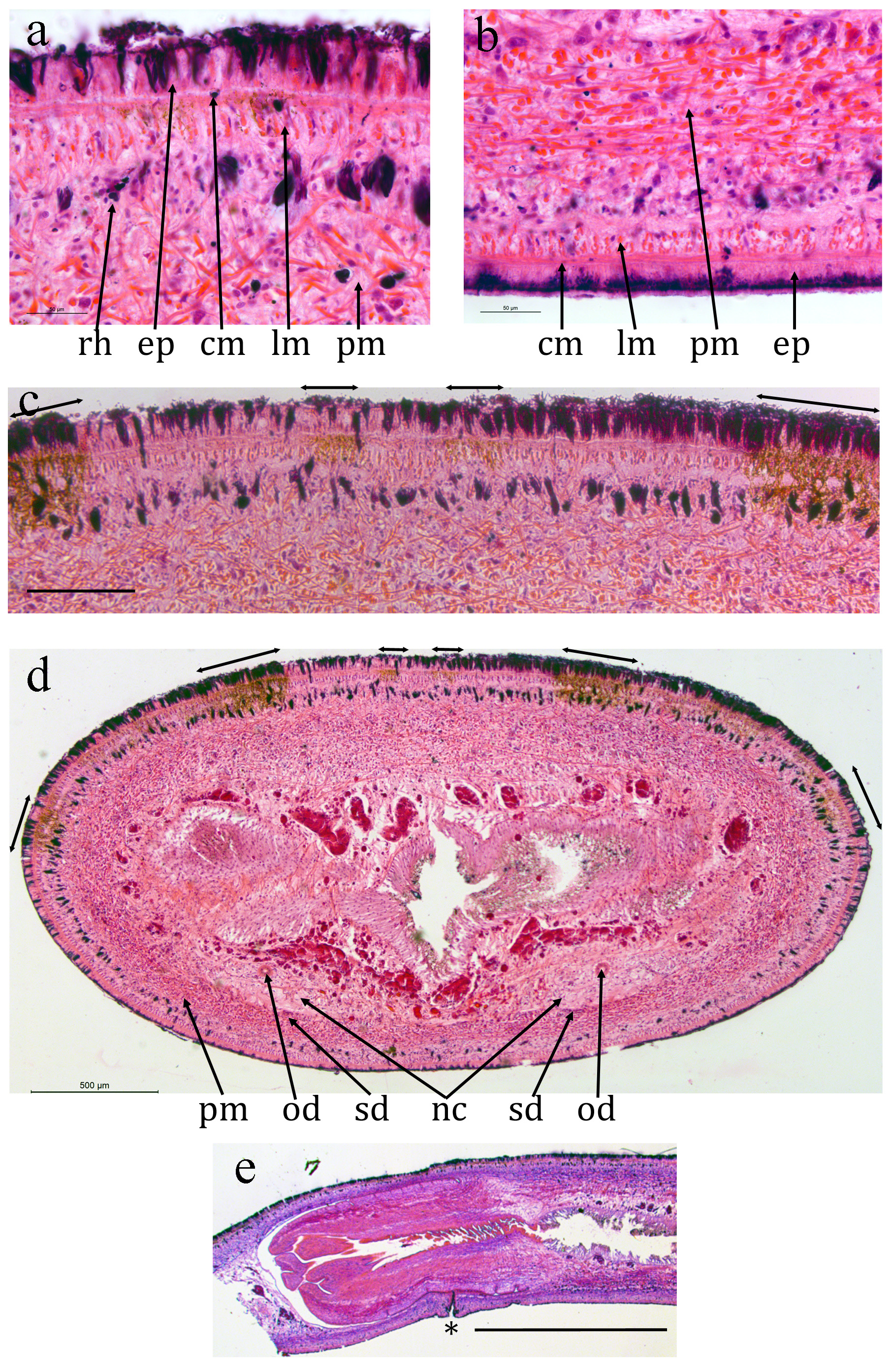
NHMUK.97 ( Figs 6 View FIGURE 6 , 7 View FIGURE 7 ): The epidermis is damaged or missing over large areas but is a monolayer of columnar cells, about 40 μm thick dorsally and about 20 μm ventrally. Many rhabdites are present both dorsally and ventrally. The creeping sole, measured from TS, is about 75% of the width.
The preserved specimen showed faint longitudinal dark lines ( Fig. 5 View FIGURE 5 ) but corresponding pigment cannot be detected in TS (it can be detected in ZMB 3506 and Cv7 — see later).
A prominent ring zone of parenchymal mixed circular and longitudinal muscle is present, about 130 μm thick ventrally and 300 μm thick dorsally.
There is a pair of ventral nerve cords with transverse commissures and a sub-epidermal muscle plexus. The nerve cords are about 1.3 mm apart.
The digestive system is typically triclad. The pharynx is cylindrical, about 6 mm long by 2 mm in diameter, the dorsal insertion a little more posterior than the ventral. The pharyngeal aperture (mouth) is about 67% along the pharyngeal pouch. The pharyngeal musculature of consists of an outer layer of circular fibres 100 μm wide, a central layer of longitudinal and radial fibres respectively about 300 μm wide and an inner layer of alternating longitudinal and circular fibres about 100 μm wide.
Numerous mature testes are present lateral to the ventral nerve cord on both sides. They are present in the TS and the LS of the pharyngeal portion, both anterior, alongside and posterior to the pharyngeal pouch, but there are none in the portion of the copulatory apparatus. Typically their maximum dimension is about 250 μm. Each is slightly elongated towards the ventral side of the nerve cord, presumed to indicate the position of the efferent duct. Sperm ducts cannot be distinguished with certainty.
Unfortunately the excised sectioned body portion is too short to show the full antero-posterior extent of the copulatory apparatus so that the entry of any sperm ducts anteriorly cannot be determined. Measured from the gonopore the male atrium is at least 7 mm long. The male atrium contains a large fold projecting from the ventral surface ( Fig. 7d, e View FIGURE 7 ).
Immature oviducts 15 μm wide with no lumen visible are present just dorsal to the nerve cord on either side. They run the full length of the sectioned portion containing the copulatory apparatus and are presumed to open into the copulatory apparatus at the posterior, female, end. Measured from the gonopore the female portion is at least 5 mm. The female atrium is simple with little differentiation.
Both male and female atria open into the gonopore with a simple opening.
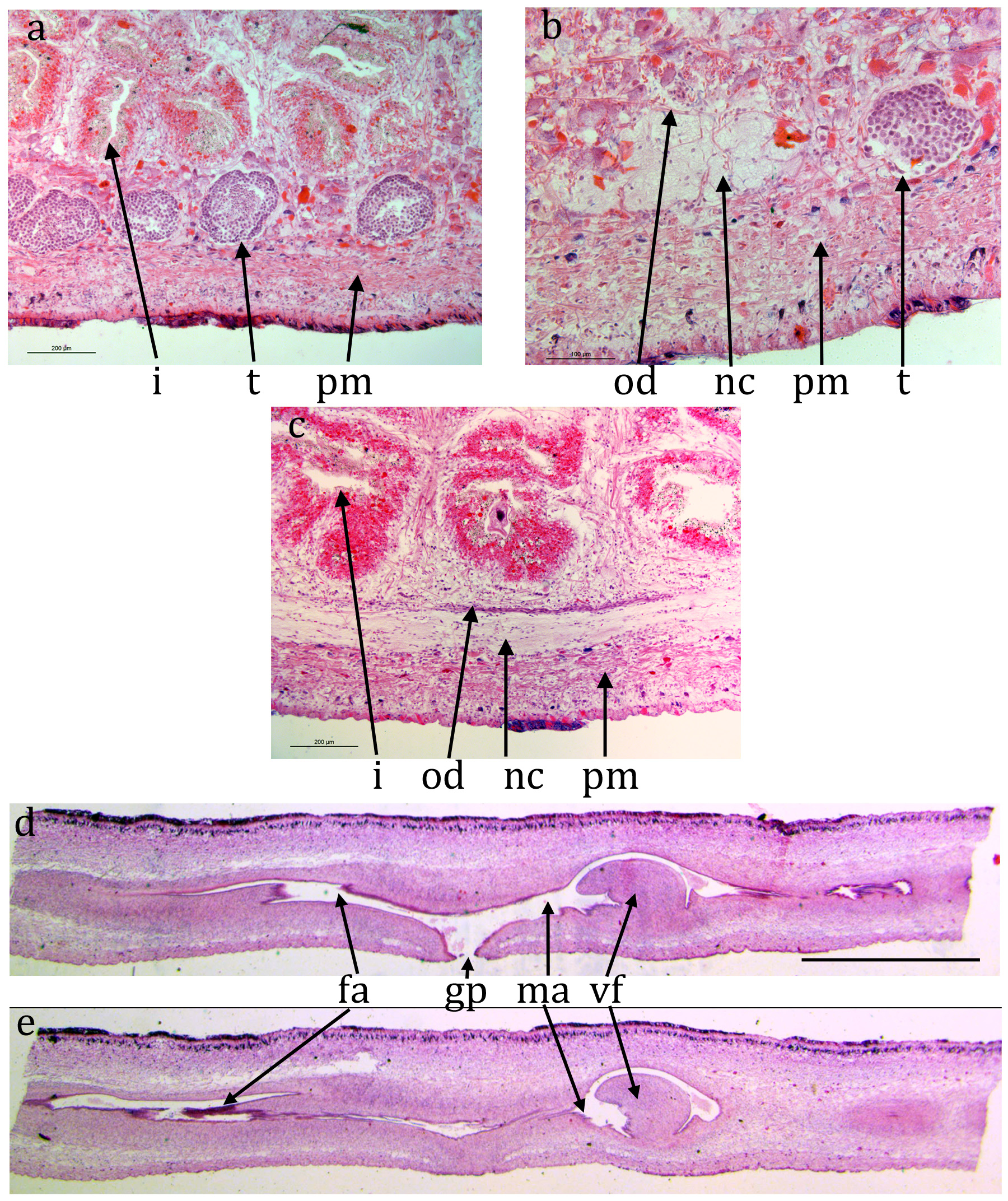
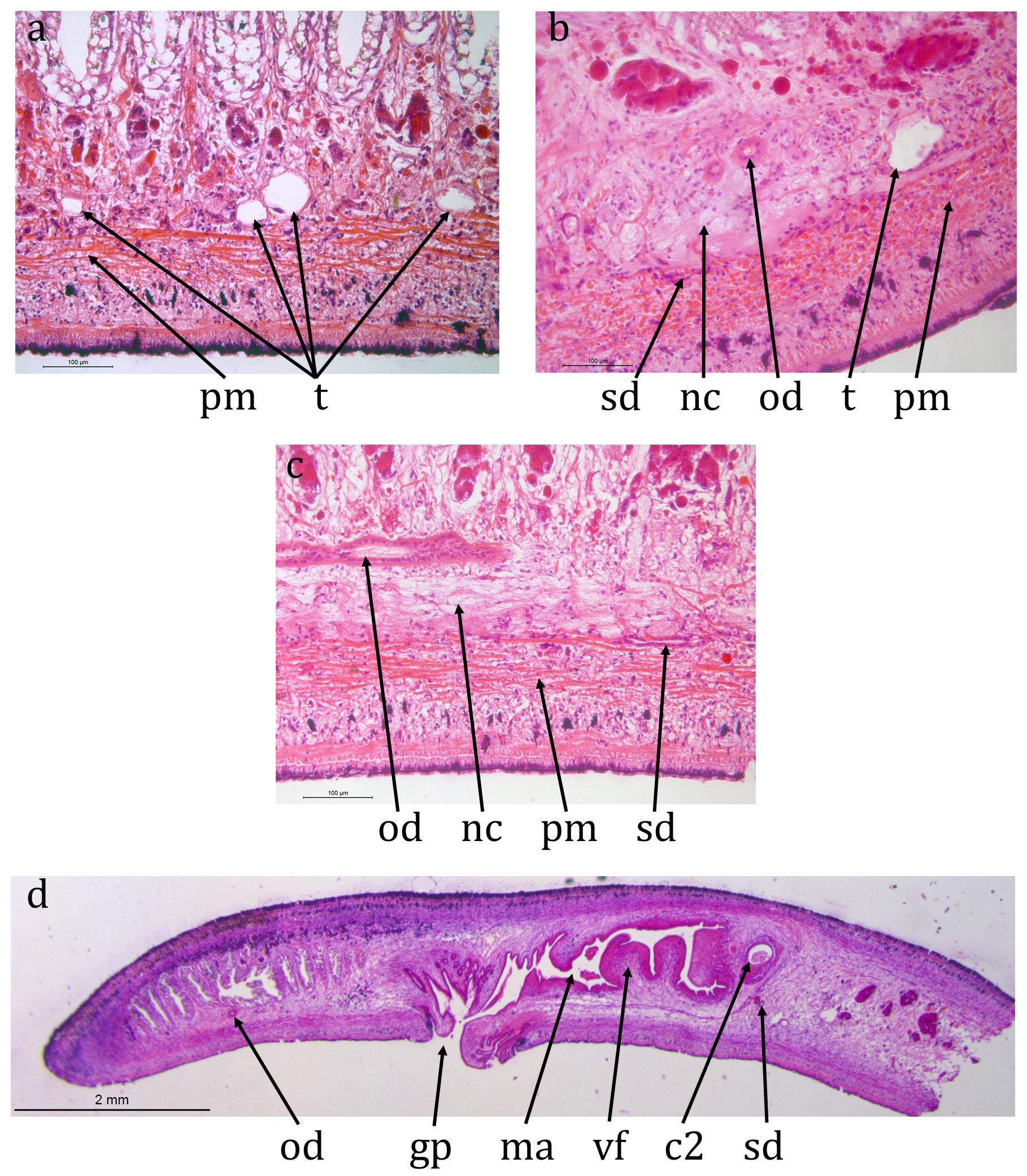
ZMB 3506 ( Figs 8 View FIGURE 8 , 9 View FIGURE 9 ): The epidermis is damaged or missing over large areas but is a monolayer of columnar cells. The creeping sole, measured from TS, is about 85% of the width.
Brown sub-epidermal pigment, probably melanic, is visible concentrated in dorsal and lateral regions ( Fig. 8c, d View FIGURE 8 ). The concentrations are: two near-medial, each about 100 μm wide and 100 μm apart, lightly pigmented and restricted to the sub-epidermal circular muscle and longitudinal muscle bundles; two dorsally but more lateral, more heavily pigmented, about 1 mm apart and about 400 μm wide and extending through the circular muscle and longitudinal muscle bundles through to the rhabditogen layer; two laterally, less heavily pigmented, each about 200 μm wide also extending through to the rhabditogen layer. The pigment concentrations are assumed to coincide with the dark stripes in living animals.
A prominent parenchymal ring zone of mixed circular and longitudinal muscle is present, about 130 μm thick ventrally and 300 μm thick dorsally.
There is a pair of ventral nerve cords with transverse commissures and a sub-epidermal muscle plexus. The nerve cords approximately 1.75 mm apart. The ventral nerve cords expand near the anterior end.
The digestive system is typically triclad. The pharynx is cylindrical, about 5 mm long by 1.7 mm, the dorsal insertion a little more posterior than the ventral insertion. The pharyngeal aperture (mouth) is about 30% along the pharyngeal pouch. The pharyngeal musculature of consists of an outer layer of circular fibres 100 μm wide, a central layer of longitudinal and radial fibres about 400 μm wide and an inner layer of alternating longitudinal and circular fibres about 200 μm wide.
Numerous immature testes, 60-70 μm in diameter, are present lateral to the nerve cord on either side. Sperm ducts cannot be discerned in TS or LS. The copulatory apparatus appears to be in the very early stages of development with little differentiation. The male part is about 4.5 mm long measured from the gonopore. At the anterior of the male part there is a sinuous narrow duct, internal diameter about 15 μm and about 500 μm long from anterior to posterior (longer when straightened) though no afferent sperm ducts are visible entering the anterior end of this duct. This duct opens into the male atrium, about 70 μm wide and 4 mm long, which is more or less linear with little differentiation.
Possible early-stage ovaries are present immediately dorsal to the ventral nerve cord on each side about 1.4 mm from the anterior end. Immature ovovitelline ducts, with no lumen visible, run posteriorly on the dorsal surface of the ventral nerve cord on either side. They can be traced posterior to the pharynx but not to the rear of the female copulatory system. No ovovitelline ducts can be seen entering the rear of the female atrium. The female atrium is about about 8.5 mm long from posterior to gonopore and 70 μm wide, also with little differentiation.
Both male and female atria open into the gonopore with a simple opening.
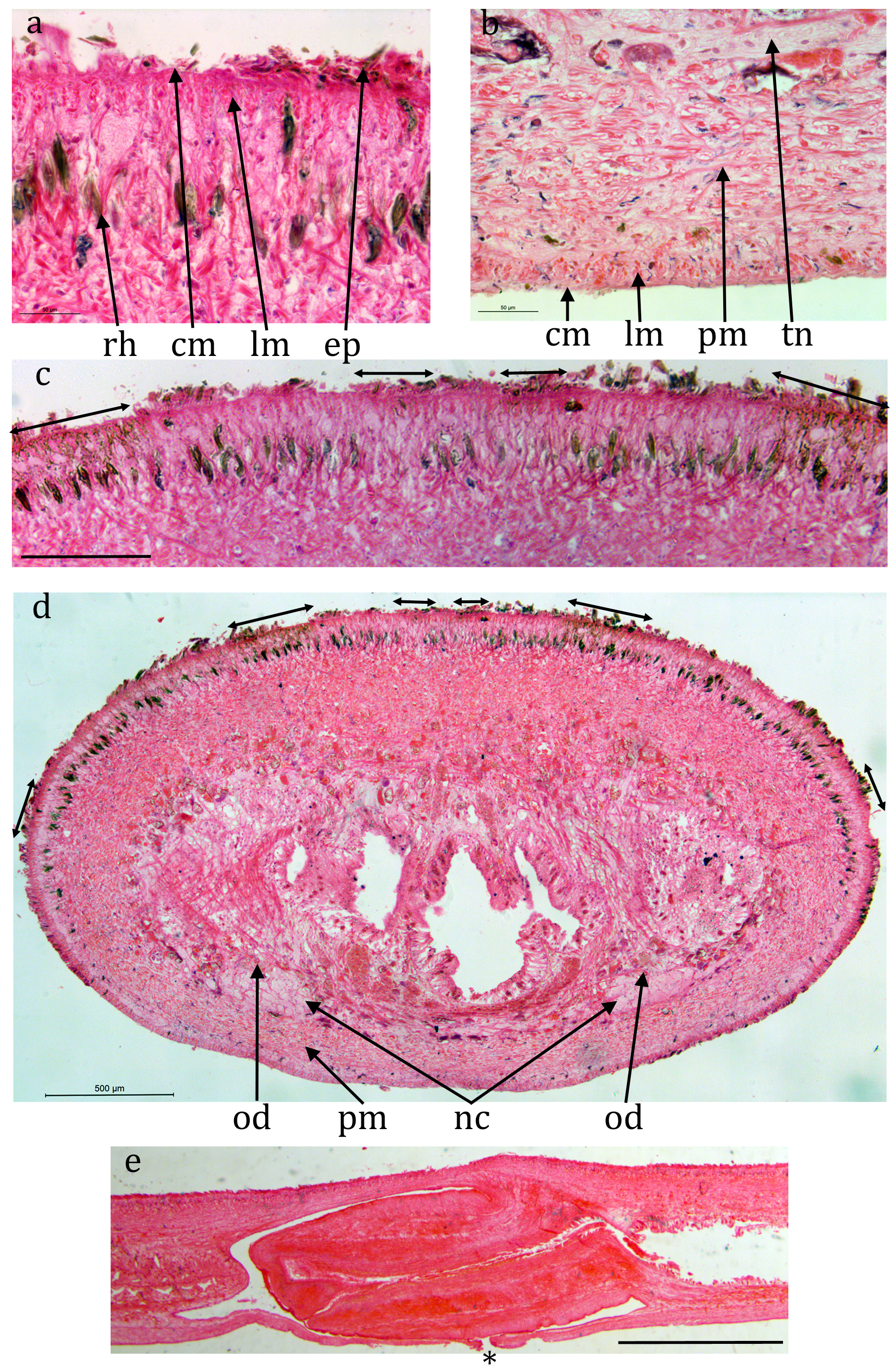
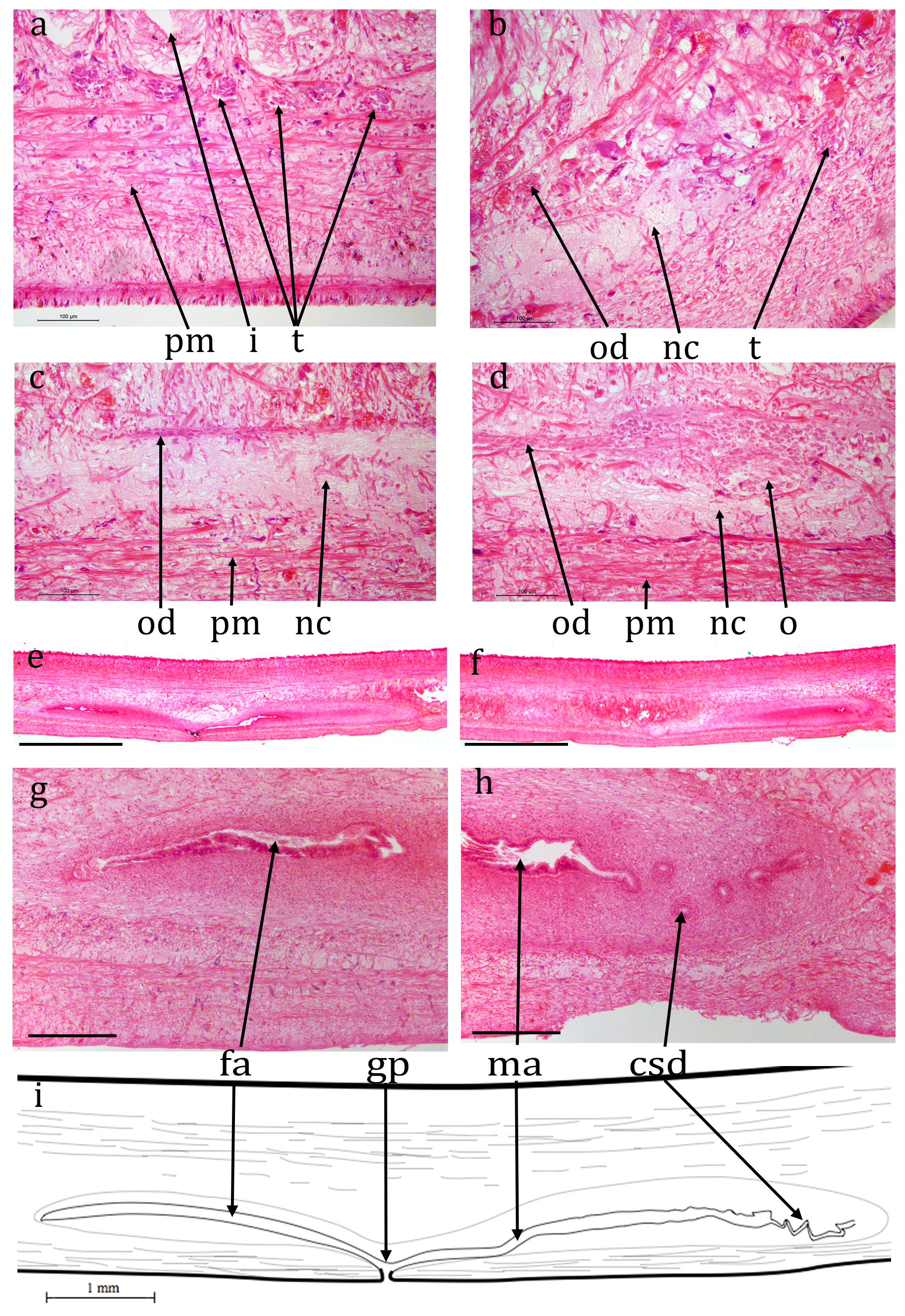
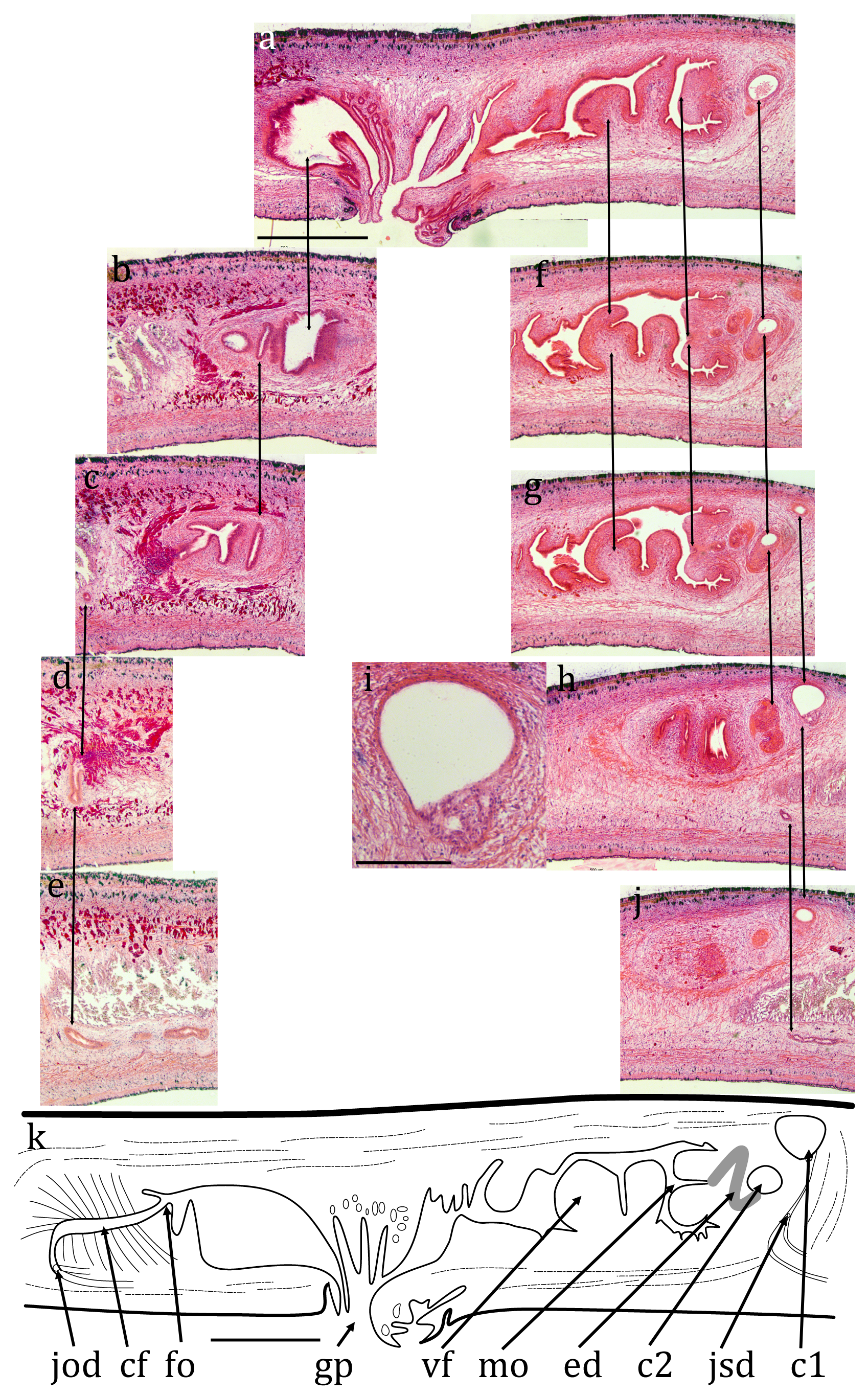
Cv7 ( NHMUK.2018.8.31.1) ( Figs 10 View FIGURE 10 , 11 View FIGURE 11 , 12 View FIGURE 12 ): The epidermis is a monolayer of columnar cells, about 50 μm thick dorsally and about 20 μm ventrally. Many rhabdites are present both dorsally and ventrally. The ventral epi- thelium is heavily ciliated, the cilia are about 5 μm long. The creeping sole, measured from transverse sections, is about 83% of the width.
Brown sub-epidermal pigment, probably melanic, is present concentrated in regions of the dorsal and lateral circumference ( Fig. 10c, d View FIGURE 10 ). The concentrations are: two near-medial, each about 100 μm wide and 100 μm apart, lightly pigmented and restricted to the sub-epidermal circular muscle and longitudinal muscle bundles; two dorsally but more lateral, more heavily pigmented, about 1 mm apart and about 400 μm wide and extending through the circular muscle and longitudinal muscle bundles through to the rhabditogen layer; two laterally, less heavily pigmented, each about 200 μm wide also extending through to the rhabditogen layer. The pigment concentrations would appear to coincide with the dark stripes in living animals.
A prominent parenchymal ring zone of mixed circular and longitudinal muscle is present, about 130 μm thick ventrally and 300 μm thick dorsally.
There is a pair of ventral nerve cords with transverse commissures and a sub-epidermal muscle plexus. The nerve cords are 1.75 mm apart.
The digestive system is typically triclad. The pharynx is cylindrical, about 3.5 mm long by about 1.5 mm, the dorsal insertion a little more posterior than the ventral insertion. The pharyngeal aperture (mouth) is about 33% along the pharyngeal pouch. The pharyngeal musculature consists of an outer layer of circular fibres 40-60 μm wide, a central layer of longitudinal and radial fibres 300 μm wide, and an inner layer of alternating longitudinal and circular fibres 100 μm wide.
No mature testes are visible in any sectioned portion. However, empty cavities are present lateral to the nerve cord on each side in the position of the testes of other specimens ( Fig. 11a, b View FIGURE 11 ) and these are interpreted as spent testes. A sperm duct is present running along the inner, medial, side of the ventral nerve cord on each side, on the dorsal margin of the parenchymal muscle ( Fig. 11b, c View FIGURE 11 ) and can be traced to the anterior of the male copulatory apparatus. Each is dorso-ventrally flattened, about 20 μm by 10 μm, the lumen is about 5 μm. No sperm are visible in any part of the sperm ducts. The sperm ducts approach the copulatory apparatus ventrally, turn dorsally, join about half way up the body, continue dorsally and open into a dorsal cavity ( Fig. 12 View FIGURE 12 , “c1”) via a complex diffuse opening ( Fig. 12j View FIGURE 12 ). This dorsal cavity is roughly circular though has an angular point ventrally, perhaps indicating an outlet though none is discernable. It measures about 350 μm anterior-posterior, by 350 μm dorso-ventral, by 440 μm laterally. Ventral and slightly posterior to this is another cavity ( Fig. 12 View FIGURE 12 , “c2”) measuring about 290 μm anterior-posterior, by 200 μm dorso-ventral, by 280 μm laterally. From the ventral side of this is what appears to be the thickened wall of a possible ejaculatory duct though no lumen is visible. This turns dorsally and then posteriorly to open into the base of a blunt projection into the broad male atrium. The atrium is about 2 mm long with prominent ventral folds ( Fig. 11d View FIGURE 11 ).
The anterior portion of the worm containing the ovaries (if present) has not been sectioned.A prominent mature ovovitelline duct ( Fig. 11b & c View FIGURE 11 ) is present dorsal to the ventral nerve cord on each side, external diameter about 50 μm, internal about 20 μm. The lumen is flagellated. The female copulatory apparatus appears to be fully developed. The ovovitelline ducts run to the posterior of the copulatory system join ventrally and the combined ovovitelline duct runs dorsally for about 250 μm, turns anteriorly and widens to form the female genital canal. The posterior part of the canal, about 400 μm long, is surrounded by copious eosinophilic glands presumed to secrete the cocoon. The anterior part has none of these glands and opens into the female atrium via what appears to be a small projection. The female atrium has some transverse folds at its posterior but broadens anteriorly before opening into the gonopore.
In the region of the gonopore, there are numerous small invaginations both dorsally and ventrally ( Fig. 12l View FIGURE 12 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Rhynchodeminae |
|
Genus |
Caenoplana variegata ( Fletcher & Hamilton, 1888 )
| Mateos, Eduardo, Riutort, Marta & Álvarez-Presas, Marta 2020 |
Caenoplana bicolor
| Vardinoyannis & Alexandrakis 2019 |
Caenoplana bicolor
| De Waart 2016 |
Caenoplana bicolor
| Sluys 2016 |
Caenoplana bicolor
| Winsor 1997 |
Australopacifica bicolor
| Ogren & Kawakatsu 1991 |
Australopacifica variegata
| Ogren & Kawakatsu 1991 |
Caenoplana bicolor
| Winsor 1991 |
Caenoplana variegata
| Winsor 1991 |
Geoplana varigata
| Winsor 1977 |
Geoplana varigata
| Winsor 1973 |
Geoplana bicolor
| Winsor 1973 |
Geoplana variegata
| Steel 1900 |
Geoplana bicolor
| Graff 1899 |
Geoplana variegata
| Graff 1899 |
Geoplana variegata
| Dendy 1894 |
Geoplana variegata
| Dendy 1893 |
Geoplana variegata
| Dendy 1892 |
Geoplana variegata
| Fletcher & Hamilton 1888 |