
Lymanopoda casneri Pyrcz & Clavijo
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4179.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:7538D34D-025A-454F-8F88-35389E9B21F7 |
|
DOI |
https://doi.org/10.5281/zenodo.5631784 |
|
persistent identifier |
https://treatment.plazi.org/id/F12C8788-FFCE-B04F-FF4D-F8C1FC52FC30 |
|
treatment provided by |
Plazi |
|
scientific name |
Lymanopoda casneri Pyrcz & Clavijo |
| status |
sp. nov. |
Lymanopoda casneri Pyrcz & Clavijo , n. sp.
( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 4, 5 View FIGURE 5 )
Type locality. Colombia, Antioquia department, municipality Belmira, vereda Río Arriba , El Morro.
Material examined. MEFLG–UN: HOLOTYPE Ƌ: municipality Belmira: pathway Los Patos towards Páramo El Morro, N 6°38.685’ W75°40.33’ 3200 m, 06.I.2012, 14.00h, BMC11146, net, A. Clavijo leg.; ALLOTYPE ♀: same locality as the holotype, N6°38.600’ W75°40.335’ 3179 m, 21.I.2012, 13.00h, BMC 14729, net, C.F. Álvarez leg., prep. genit. 01/ 26.IV.2013, J. Lorenc, MZUJ GoogleMaps ; Paratypes (13 ƋƋ and 1 ♀): same locality as the holotype: 1 Ƌ: 3234 m, 21.I.2012, 12.50h, BMC 14728, net, C.F. Álvarez leg. GoogleMaps ; 1 Ƌ: 3244 m, 23.I.2012, 11.50h, BMC14677, net, C.F. Alvarez leg.; 2 Ƌ: 3226 m, 11.V.2012, 12.50 and 13.30h, BMC15116 and BMC15119, net, A. Clavijo leg.; 1 Ƌ: 3220 m, 12.V.2012, 10.15h, BMC16521, net, C.F. Álvarez leg.; 3 ƋƋ and 1 ♀: 3193 m, 14.V.2012, 10.45h, BMC15203, BMC15209 prep. molec. 25/ 23.III.2013 A. Zubek MZUJ, BMC 15216 in Chusquea sp. prep. molec. 26/ 23.III.2013 A. Zubek MZUJ, BMC15205 eggs photo, net, J. Duque and C.F. Álvarez leg.; 1 Ƌ: 3227 m, 15.V.2012, 12.00h, BMC16269, net, A. Clavijo leg; RCCP: 1 Ƌ: Páramo de Belmira , 3100 m, 05.I.2011, i789, C. Prieto leg.; 1 Ƌ: same data, i790, C. Prieto leg. GoogleMaps ; 1 Ƌ: same data, i802, C. Prieto leg. GoogleMaps ; MZUJ: 1 Ƌ: same locality as the holotype, 3226 m, 11.V.2012, 13.30h, BMC 15112, net, A. Clavijo leg. GoogleMaps
Diagnosis. From among several predominantly white species of the genus Lymanopoda Westwood, 1851 , L. casneri n. sp. most closely resembles externally L. zebra Pyrcz & Rodríguez, 2007 , which has HWD black venal stripes, and L. vivienteni (Apolinar) , the latter however has a much more pointed FW apex. Contrary to L. zebra Pyrcz & Rodriguez 2007 in L. casneri most of median area of FWD is free of black.
Description. MALE ( Fig. 2 View FIGURE 2 A): Head: Eyes dark brown, covered with long, black hair; antennae half the length of costa, slender, chestnut, with white scales at base of each flagellomere, club darker brown, two times the width of shaft, made of 9–10 segments, spoon–like. Thorax: black, covered with sparse black and white hair, dense grey on protothorax; legs dirty yellow, covered with chestnut, black and sandy yellow scales, thick on tibia and tarsus, sparse on femur. Abdomen: black, dorsally and laterally covered with black hair, ventrally with white hair and scales. Wings: FW (length: 20–23.5 mm, mean: 21.5, n=3) elongated with an acute apex, straight outer margin and a gently curved tornus; FW fringes white, grey and black, except at tornus were only white. HW fringes mostly white. FWD: Basal and postbasal area, and along anal margin blue gray; discal cell mostly black except for a mid– cell white patch; postdiscal area to outer margin black from costa to vein M3 except for three elongated postdiscal patches from R5–M1, M1–M2 to M2–M3, and three smaller, white subapical streaks in the same cells; two large black rounded patches in mid cell M3–Cu1 and Cu1–Cu2; a black submarginal to marginal area spreading to outer margin along veins Cu1 and Cu2 leaving free two white triangular patches pointing distally in M3–Cu1 and Cu1– Cu2. FWV: Ground colour white; sandy yellow along costa, along outer margin to apex, and along outer margin were four to five marginal, triangular patches are suffused to a varied degree with sandy yellow, additionally all the veins are marked with sandy yellow; a roughly rectangular black mid discal cell patch with a diffused inner margin gradually transforming into a light grey basal suffusion; a larger rectangular patch extending over discal cell distal extremity into postdiscal area from subcostal to vein M3, two minute subapical black dots and two larger rounded postdiscal spots in M3–Cu1 and Cu1–Cu2, all black elements of the colour pattern are edged with a light gray suffusion which reflects the black area on the upperside. HWD: Basal to mid discal cell and from vein Cu1 to anal margin blue gray; most median, postmedian and submarginal area white; black triangular patches gradually widening along all the veins from postdiscal area to outer margin. HWV: Mostly sandy yellow, except for white intravenous stripes, the largest of which is the one which extends from wing base along discal cell and cell M2–M3 to outer margin.
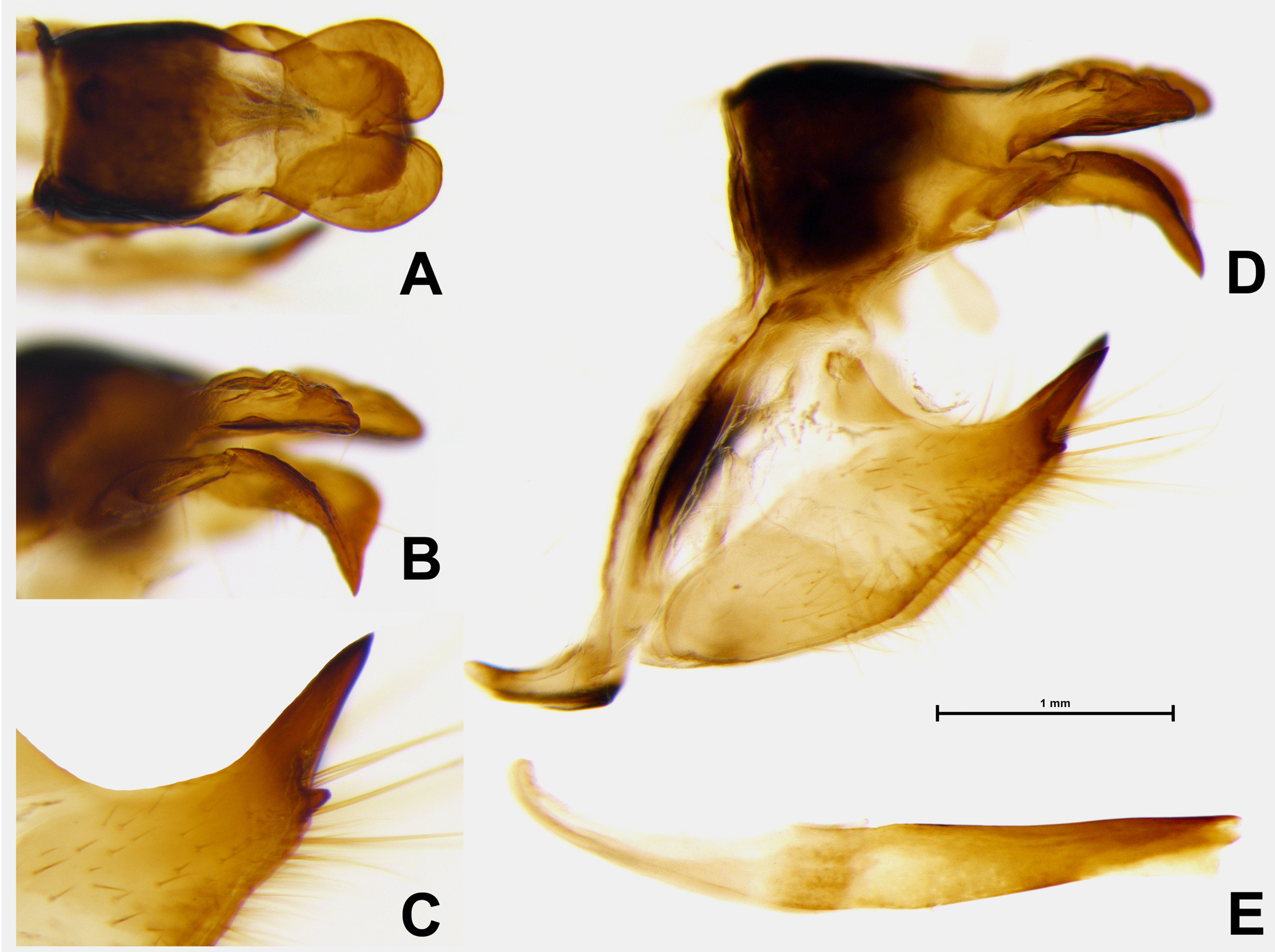
MALE GENITALIA ( Fig. 3 View FIGURE 3 ): The new species shows some diagnostic characters of male genitalia which separate it immediately from any other predominantly white patterned paramo dwelling Lymanopoda . In particular, the bifurcate superuncus is very long, nearly as long as the uncus, which is a character frequent in the species of L. obsoleta clade (Pyrcz et al. 2011) but in L. huilana Weymer, 1980 L. tolima Weymer, 1911 , L. melia Weymer, 1912 and L. zebra it is short or very short, never exceeding one–third the length of uncus. The uncus is also very particular, instead of being stout, flattened laterally and swollen in the middle, such as the uncus of the above mentioned species, it is sledge–like, that is considerably flattened dorso–ventrally with a wide median bruise. The subuncus is very small, nearly completely atrophied whereas in other white paramo species it is adhered to the base of uncus but rather well developed. The subscaphium is membraneous, without any sclerotization at all, compared to the weakly sclerotized homologous structure of other white paramo Lymanopoda found in the Central Cordillera , or the heavily sclerotized subscaphium of two Eastern Cordillera species, L. mirabilis Staudinger, 1897 and L. vivienteni (Apolinar, 1924) , and indeed of most other congeners. The valva is shaped as in L. melia , that is with a massive basal half, tapering to a short but pointed apical extremity. However in L. melia , the apical part ends in two sharp but short processes of about the same length whereas in the new species it terminates into a long and heavily sclerotized sharp process, coupled with a short, ventral process. The saccus and the aedeagus is similar to other white Central Cordillera paramo Lymanopoda species.
FEMALE ( Fig. 2 View FIGURE 2 B): Sexual dimorphism slight, lighter and paler on both dorsal and ventral wing surface, with a more uniform, almost uniform light beige HWV; FW length 22–23 mm.
FEMALE GENITALIA ( Fig. 4): Papillae anales large, nearly the size of bursa copulatrix, rounded in lateral view, with a rather well sclerotized basal half, and a membranous distal half covered with dense but delicate bristles, anterior apophysis developed, roughly two–thirds the length of papillae from base to apex; lamella antevaginalis large, heavily sclerotized, slat like; lamella postvaginalis large, less sclerotized, bowl shaped; ductus bursae wide, tubular, approximately as long as the length of bursa copulatrix, sclerotized in basal half, abruptly opening into bursa copulatrix, with ductus seminalis originating at its middle; bursa copulatrix oval, without any visible signa.
EGGS ( Fig.5 View FIGURE 5 ): Eggs presented a translucent white colour, are oval shaped with longitudinally aligned stripes and micropyle area is rough with circular shapes.
Etymology. This species is dedicated to the American entomologist, Kayce Casner, who has significantly contributed to the knowledge in the genus Lymanopoda .
Comments. Morphological and genetical evidence yield different results. On the one hand, despite the similarity of colour patterns, genital morphology strongly indicates that L. casneri does not belong to the species of the “white group” found in in the Colombian Central Cordillera or Western, such as L. zebra , L. melia , L. huilana and L. tolima , nor to the two white paramo species occurring in the Eastern Cordillera , L. vivienteni and L. mirabilis . It appears that the white colours and elongated wings is a simple convergence to the paramo habitat among the unrelated species of the genus Lymanopoda . Male genitalia of the new species present a series of strong synapomorphous character with L. hazelana Brown, 1943 found in central and southern Ecuador and northernmost Peru. On the other hand, barcode data while supporting the separation of the new species indicate its closer a genetic affinity with the “white” L. tolima than with L. hazelana expressed in genetic distances of 3.5% with L. tolima and 8% with L. hazelana (Appendix 1) ( Fig. 15-16 View FIGURE 15 View FIGURE 16 ). However, considered the molecular data used here, this has to be considered with due reservation, whereas available morphological evidence is more comprehensive.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |