
Batillipes dandarae, Santos, Erika, Da Rocha, Clélia M. C., Jr, Edivaldo Gomes & Fontoura, Paulo, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4243.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:E3A1C72B-EB8F-4EE0-9BF7-99757331E193 |
|
DOI |
https://doi.org/10.5281/zenodo.5617030 |
|
persistent identifier |
https://treatment.plazi.org/id/F1778B6A-5B4B-FFEC-63BD-05541977FC83 |
|
treatment provided by |
Plazi |
|
scientific name |
Batillipes dandarae |
| status |
sp. nov. |
Batillipes dandarae View in CoL sp. nov.
( Figs 4–5 View FIGURE 4 View FIGURE 5 ; Table 2 View TABLE 2 )
Diagnosis. Medium sized Batillipes with cylindrical undivided primary clavae. Papillar secondary clavae present. Sensorial leg spines and cirri, including all cephalic cirri, lateral cirri and cirri E, with an optically dense dot near their blunt tips. Sensorial spine on leg I inserted on the posterior part of the leg and turning frontwards. Toes 3 and 4 of legs IV with different lengths. Lateral processes between legs absent. Pointed triangular caudal appendage very variable in shape and size among specimens. Finely punctated cuticle comprised of small pillars; dorsal punctations uniformly distributed. Rosette-like female gonopore separated from the anus by a groove. Males with circular gonopore with crescent-shaped cuticular fold. Anus surrounded by a peculiar cuticular structure constituted by six platelets.
Type locality. Gunga Beach , Alagoas, Brazil ( 9°51'45"S, 35°54'17"W). GoogleMaps
Type material. Holotype: adult, female (slide CVII-81) collected at Gunga Beach, mounted in glycerol. Allotypic male (slide TARD/UFRPE 02-18), collected at Forte Orange Beach, mounted in polyvinyl alcohol. Paratypes: 23 specimens (6 females, 17 juveniles) collected at Forte Orange Beach, mounted in polyvinyl alcohol (slides TARD/UFRPE 02-15 to 02-21); 1 female collected in Ponta do Sal Beach, mounted in polyvinyl alcohol (slide CVII-83); 2 females collected in Patacho Beach, mounted in glycerol (slide TARD/UFRPE 02-22).
Type repository. The type material ( holotype slide CVII-81 and paratype CVII-83) is deposited in the collection of the Department of Biology, Faculty of Sciences , University of Porto , Portugal, and the other slides in the collection of Tardigrades—UFRPE ( Laboratory of Meiofauna , Department of Biology , Universidade Federal Rural de Pernambuco, Brazil).
Etymology. The specific name, dandarae , is in honor of “Dandara dos Palmares”, an ex-slave warrior, who led the fight against slavery in Brazil in the XVII century.
Ecological note. B. dandarae sp. nov. was mainly found in shallow sublittoral fine to medium calcareous and quartz sands in low energy and estuarine beaches as Gunga Beach and Forte Orange Beach and occasionally in high energy beaches (Ponta do Sal) and in reef pools as Patacho Beach.
Description of holotype. Female, 194 µm long (204 µm including the caudal apparatus) and 68.9 µm wide between the third and fourth pairs of legs ( Fig. 4 View FIGURE 4 A, B). Eyes present (very faint and difficult to see, fig 4 D, probably dissolved by effect of the mounting media). Trapezoid head with eleven cephalic appendages ( Figs 4 View FIGURE 4 D, E, 5 A): Internal cirri inserted dorsally on the frontal edge of the head are 23.2 µm long, bearing cirrophores (about 3.1 µm long). External cirri 16.1 µm long ( Fig. 4 View FIGURE 4 E), with indistinct cirrophores, inserted ventro-laterally, ventral to the pedestals bearing lateral cirri and primary clavae. The median cirrus, with cirrophore (about 3.4 µm long), is 24.8 µm long ( Fig 4 View FIGURE 4 D). The lateral cirrus, 28.2 µm long, is located dorsally in relation to the unconstricted tubular primary clava which is 13.3 µm long. These two appendages share a common pedestal ( Fig 5 View FIGURE 5 A). A van der Land’s organ is present at the base of the primary clava. All the cephalic cirri, including external cirri, and lateral cirri have an optically dense dot (black dot when observed under PHC) near their blunt tips. Indentation or notch in the frontal margin of the head between the external cirrus and the pedestal bearing the primary clava and lateral cirrus not observed. In the frontal edge of the head well developed (major diameter about 6.8 µm) secondary clavae are present ( Fig 5 View FIGURE 5 A).
Ovoid pharyngeal bulb 19.5 µm long and 19.0 µm wide. Placoids not visible after slide mounting. Mouth opening located at the top of an ovoid buccal cone (12.1 µm long and 18.2 µm wide) about 10 µm distant from the frontal edge ( Fig. 4 View FIGURE 4 E).
A dorsal blunt enlargement is visible in the scapular region at the level of legs I Figs 4 View FIGURE 4 A, B, 5 A). No other body lateral projections or cuticular lateral processes are present. The caudal apparatus is constituted by a cuticular conical-shaped projection, 8.9 µm long.
Sensorial spines present on all legs. Sensorial spine on leg I is the shortest, about the same length on legs II and III, and longer on legs IV (9.4; 11.3; 11.2, and 12.7 µm long, respectively). Leg I sense organs are not subdivided into different parts. They are similar in shape to sense organs on legs II and III that are spine-like. However, leg I sense organs are inserted on the posterior part of the leg and revolved frontwards ( Fig. 5 View FIGURE 5 B). All the sense organs on legs I–III have an optically dense dot near their blunt tips. The sense organ on leg IV is divided into a cirrophore (1.3 µm long), a proximal portion (4.7 µm long) and a distal portion (6.4 µm long) with a basal van der Land’s organ and an optically dense dot near the tip. Dorsal cirri E are blunt and short (18.7 µm long) with no evident cirrophores and also with a dot near the tip.
Telescopic legs without tibial papillae. Toes with distal stalks considerably enlarged (2.6 µm wide), terminated by small ovoid suction discs (3.7 µm long and 3.4 µm wide on leg IV) with conspicuous braces and well developed peripheral thickenings. In the first three pair of legs, toe 2 is the shortest, toes 3 and 5 are the longest and toes 1, 4 and 6 are medium sized (13.4, 4.7, 16.2, 10.2, 17.9 and 12.7 µm long, respectively for toes 1 to 6 of leg II). In the fourth pair of legs ( Figs 4 View FIGURE 4 A, B, 5 C) toes conform to the pattern of the D group of species proposed by Kristensen and Mackness (2000), with toes 3 and 4 of different lengths (10.1 and 12.7 µm long respectively). In legs IV toes 1 and 6 are of intermediate size and similar in length; toes 2 and especially toes 5 are the longest (this character was observed in paratypes; in the holotype only toe 1, 14.4 µm, was measurable).
Dorsal cuticle uniformly and finely punctated, with about 15 pillars/10 µm, comprised of short pillars ( ca. 1 µm high) with some delicate transverse lines (about eight) and without smooth regions. Less dense punctation on ventral cuticle which exhibit some transverse folds.
Gonopore rosette-shaped ( Figs 4 View FIGURE 4 C, 5C) separated from the anus by a very small groove. The anus, 7.3 µm posterior from the gonopore, is surrounded by a peculiar cuticular round structure, constituted by six platelets.
Remarks. Sexual dimorphism was not evident in the only male (138 µm long) found. This male is similar to females in both qualitative and metric characters (except for their circular gonopore with crescent shaped cuticular fold, distant 6.5 µm from the anus). Juveniles, with six toes on each leg but without a visible gonopore, were also similar to adults. Four-toed larvae were not found. Sense organ on legs I inserted on the posterior part of the leg and turning frontwards, similar to the one described for B. noerrevangi Kristensen, 1978 is consistently present in all the examined specimens. Shape and size of the caudal apparatus are strongly variable among individuals.
Differential diagnosis. Batillipes dandarae sp. nov. in having medial toes 3 and 4 on leg IV of different lengths and respectively different from toe 1 and toe 2, belongs to the D group of species ( Kristensen and Mackness 2000). Only six known species, including B. brasiliensis sp. nov. described above, exhibit this toe pattern on leg IV. However, similarly to B. dandarae sp. nov., only two other species included in this D group, B. africanus and B. tubernatis , lack body cuticular lateral processes.
B. tubernatis View in CoL was originally described by Pollock (1971) as having a rounded and swollen caudal contour and not a real caudal appendage. In the emended description ( McKirdy 1975), some specimens of B. tubernatis View in CoL can exhibit a “strong, single, swollen-based caudal spike inserting as a continuous extension of the body”, similar to the caudal apparatus of the new species. However, in Batillipes dandarae View in CoL sp. nov. the caudal appendage is present in all the specimens, including young. Moreover, the new species differs from B. tubernatis View in CoL in the aspect of dorsal punctations: they are uniformly distributed and finely punctate in the new species, while apunctate areas and different sized punctations occur in B. tubernatis View in CoL . In B. dandarae View in CoL sp. nov., and contrary to B. tubernatis View in CoL , the secondary clavae are well developed, protruding from the frontal head border. In addition, according to McKirdy (1975), in B. tubernatis View in CoL toe suction discs are rounded-quadrate to subcircular in shape with slightly indented frontal edge (see Fig. 12 B in McKirdy 1975), while they are ovoid in the new species ( Fig. 5 View FIGURE 5 A, C).
Batillipes dandarae View in CoL sp. nov. and B. africanus View in CoL are very similar each other. In B. dandarae View in CoL sp. nov., compared to B. africanus View in CoL , pedestals bearing primary clavae and lateral cirri are less pronounced. Despite the variability in shape and size, the caudal apparatus is more consistently triangular shaped in B. dandarae View in CoL sp. nov., and never in the shape of an elongated spine as sometimes occurs in B. africanus View in CoL . On the other hand, specimens of the new species without caudal apparatus were not found. Segmental folds are weakly marked in the new species, preventing the differentiation of body sections, contrary to B. africanus View in CoL where a cephalic, a caudal and three body sections are evident, including in first stage larvae. Another difference between the two species refers to tibial papillae that are present in B. africanus View in CoL only. In addition, the following characters, by their taxonomic relevance, deserve special attention. i) Shape of the scapular region: in B. dandarae View in CoL sp. nov. a conspicuous dorsal blunt enlargement is present in the scapular region at the level of legs I, while B. africanus View in CoL lacks this enlargement and the body is more rectangular-shaped. ii) Morphology of leg I sense organs, that are spine-like, inserted in the posterior region of the legs and revolved frontwards in the new species and not articulated, straight and parallel to the toes, as in B. africanus View in CoL . In addition, in the new species sense organs on legs III are longer, about twice the length, than those of B. africanus View in CoL (range 10.0–11.2 µm in the new species; 5.0–6.0 µm in B. africanus View in CoL ). iii) Structure and morphology of the gonopore and the anus: in the new species the female gonopore has the typical rosette-like shape without additional structures or special cuticular areas and the anus is surrounded by a round structure constituted by six cuticular platelets. In B. africanus View in CoL the female gonopore has two posterior apunctated areas and the anus is covered by three platelets.
Small dots near the tips of sensorial structures, appearing as black dots (optically dense) under phase contrast microscopy, present in B. dandarae sp. nov. were not described for both B. tubernatis and B. africanus . Optically dense regions below the tip of cephalic cirri were observed by Gallo D’Addabbo et al. (2000), redescribing B. dicrocercus . Probably, the observation of these dots had been neglected in the description of other Batillipes species; therefore this character should be carefully used in comparisons of different species.
As previously mentioned, the structure of the revolved sense organ on leg I of B. dandarae sp. nov. is shared with B. noerrevangi . The two species cannot be confused because they exhibit a different toe pattern ( B. noerrevangi belongs to the A group of species proposed by Kristensen and Mackness 2000, which have the middle toes of legs IV of equal length). Both species can be found in low saline waters. The new species occurring mainly in estuarine beaches, as it is the case of Forte Orange Beach that, according da Rocha et al. (2004), has a salinity value of 5–29 PSU, while B. noerrevangi can be found in low saline waters of the North Sea (4–20 PSU) and the Baltic Sea (3–11 PSU) ( Kristensen 1978, Zawierucha et al. 2015).
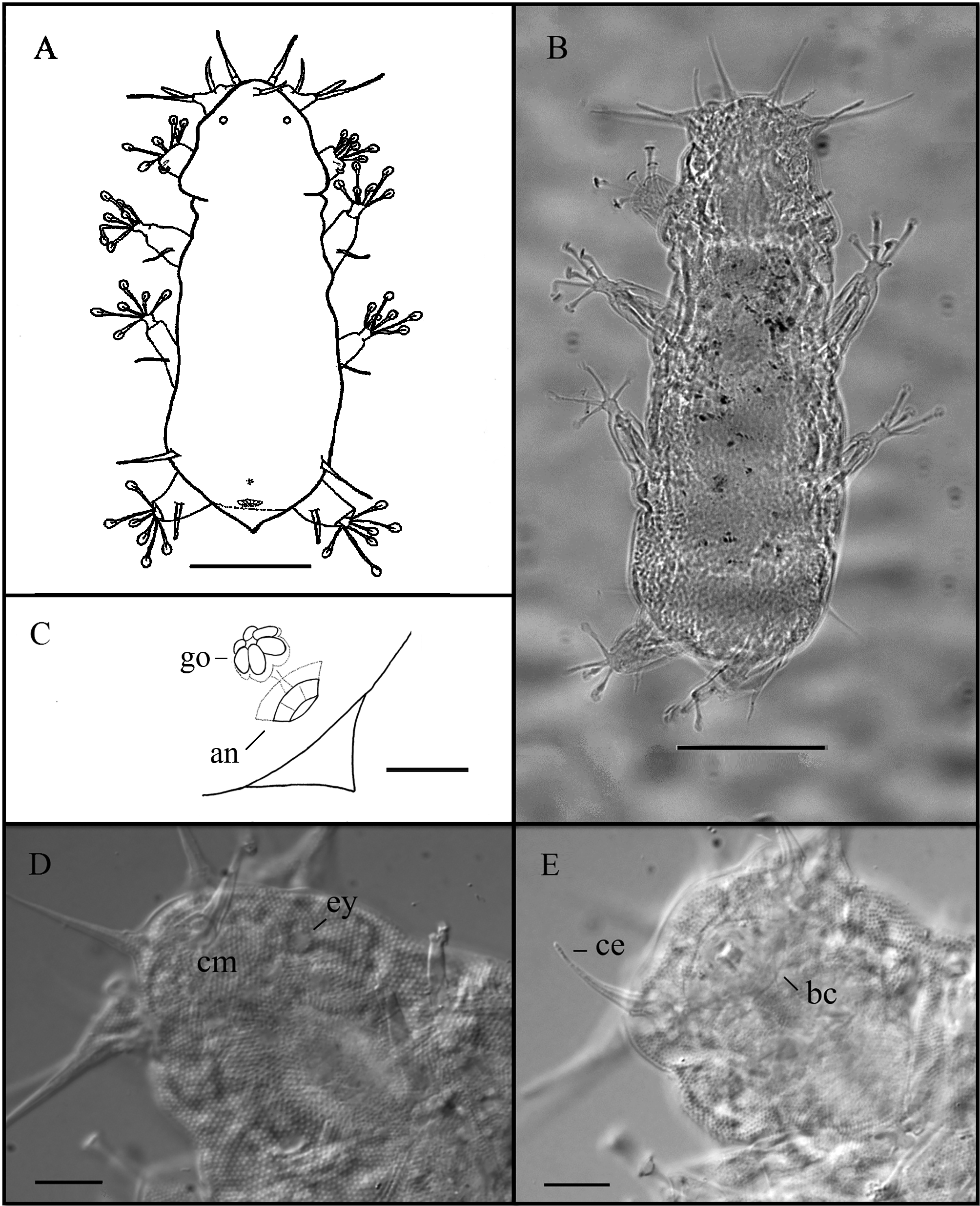
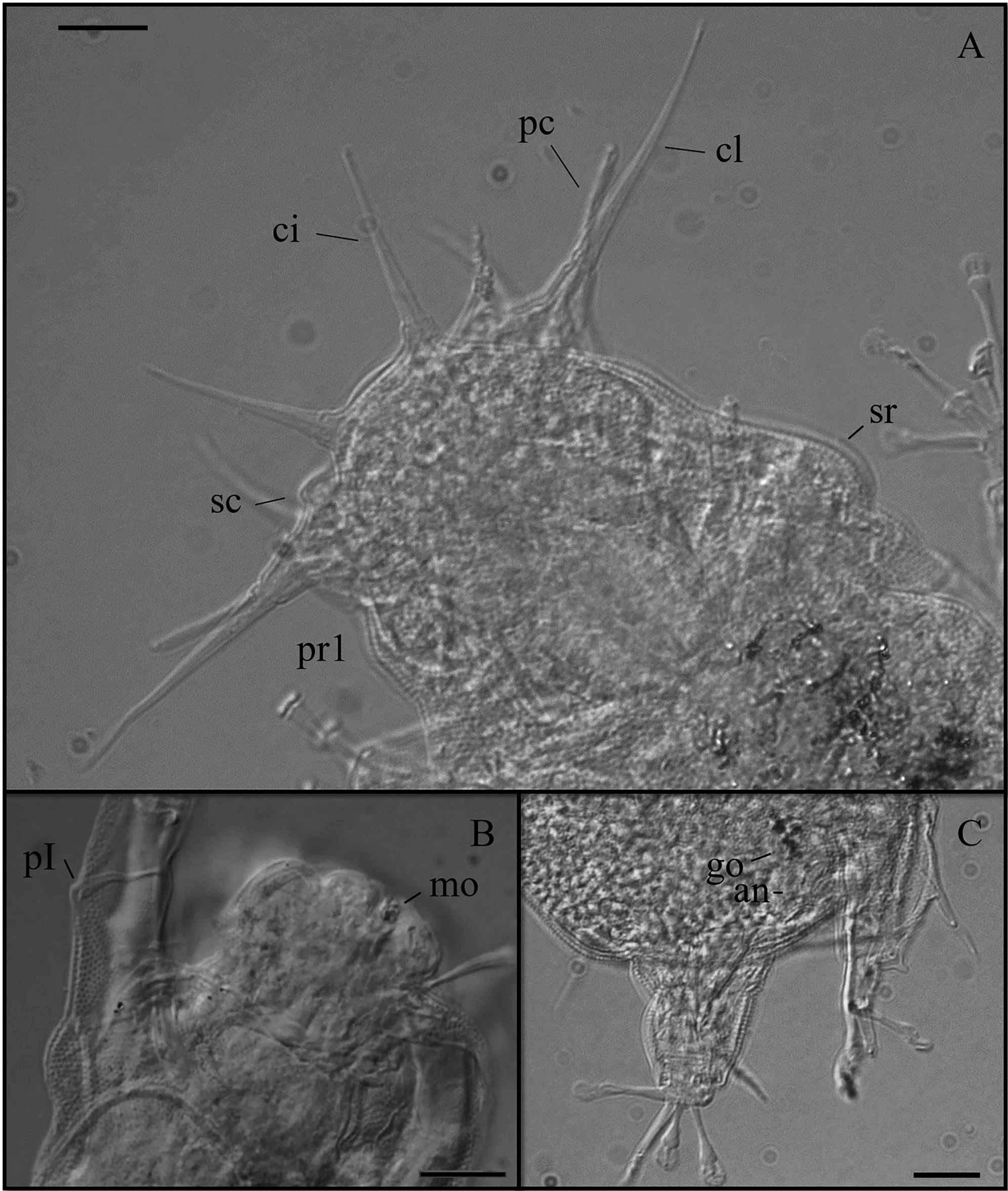
TABLE 2. Measurements (in µm) of selected morphological structures of specimens of Batillipes dandarae sp. nov. (Holo—Holotype; SD—Standard deviation; Range refers to the smallest and largest measured specimen / structure; Nnumber of specimens / structures measured; Allo—Allotype).
| FEMALES adults Holo Mean ± SD (Range); N | MALE adult Allo | JUVENILES 6-toed Mean ± SD (Range); N | |
|---|---|---|---|
| STRUCTURES Body length (BL) BL without caudal apparatus | 204 176.5 ±36.1 (129.7–224.8); 8 194 165.4 ±34.6 (119–208); 10 | 137.5 124.4 | 167.9 ±33.96 (127.4–219.1); 12 155.8 ±27.38 (122.2–200.1); 15 |
| Body width Median cirrus Internal cirri | 68.9 53.5 ±14.4 (33.6–73.9); 9 24.8 19.9 ±3.4 (13.5–24.8); 7 23.2 19.1 ±2.9 (14.3–23.2); 10 | 56.5 15.1 15.3 | 53.2 ±10.95 (37.3–71.4); 9 19.4 ±4.29 (14.2–27.7); 9 18.2 ±2.51 (13.5–21.6); 11 |
| External cirri Lateral cirri A Primary clavae | 16.1 13.4 ±2.8 (9.7–17.3); 9 28.2 25.9 ±5.1 (18.2–33.8); 10 13.3 11.0 ±2.03 (6.7–14.4); 10 | 8.6 19.6 8.7 | 14.3 ±3.36 (9.1–21.3); 10 23.3 ±2.80 (19.5–28.1); 11 11.1 ±1.78 (7.8–14.0); 11 |
| Primary clava / cirrus A Secondary clavae Cirri E | 0.47 0.43 ±0.05 (0.35–0.51); 10 6.8 6.7 ±1.06 (5.2–8.4); 8 18.7 15.8 ±2.9 (11.1–18.7); 6 | 0.44 5.8 9.8 | 0.47 ±0.04 (0.38–0.54); 11 5.5 ±1.20 (3.6–7.7); 11 13.4 ±4.93 (7.7–24.2); 9 |
| Leg I spine Leg II spine Leg III spine | 9.4 6.7 ±2.83 (3.4–10.8); 8 11.3 8.6 ±2.37 (4.1–11.3); 6 11.2 10.6 ±0.49 (10.0–11.2); 4 | ??? | 7.5 ±3.21 (4.6–12); 4 6.0 ±1.66 (4.5–8.8); 5 7.0 ±0.86 (6.2–7.9); 4 |
| Leg IV sense organ Leg IV base Leg IV spine | 12.4 10.1 ±1.72 (7.1–12.7); 6 5.5 4.4 ±0.66 (3.6–5.5); 8 4.6 4.8 ±0.96 (3.6–6.7); 8 | 8.6 3.4 5.2 | 10.3 ±2 (6.9–14.1); 11 4.2 ±0.97 (2.8–6); 8 5.0 ±1.01 (3.7–6.8); 8 |
| Caudal apparatus Legs I–III Toe 1 Toe 2 | 8.9 12.0 ±4.32 (6.2–19.3); 7 13.4 9.6 ±2.40 (6.3–13.4); 5 4.7 5.5 ±0.84 (4.5–6.7); 5 | 13.1 7.3 4.0 | 11.9 ±5.59 (1.7–22.0); 12 8.3 ±1.61 (5.7–11.3); 9 4.3 ±1.10 (3.1–6.3); 9 |
| Toe 3 Toe 4 Toe 5 | 16.2 12.6 ±3.20 (7.2–16.2); 5 10.2 9.7 ±1.23 (8.2–11.3); 5 17.9 15.9 ±2.89 (10.3–17.9); 5 | 11.6 6.1 15.0 | 12.3 ±2.55 (8.1–16.9); 9 8.9 ±1.02 (7.2–10.6); 9 16.3 ±2.15 (14.3–20.0); 9 |
| Toe 6 Leg IV Toe 1 Toe 2 | 12.7 11.3 ±1.53 (8.8–12.7); 5 14.4 13.0 ±1.96 (10.0–15.6); 6? 16.6 ±3.61 (12.5–22.5); 6 | 9.0 11.6 16.0 | 9.6 ±1.59 (7.5–12.9); 9 11.7 ±1.48 (10.1–14.5); 8 15.8 ±2.38 (13.2–19.5); 8 |
| Toe 3 Toe 4 Toe 5 | 10.1 9.2 ±1.60 (6.8–12.3); 7 12.7 12.3 ±1.83 (10.0–16.0); 7? 17.6 ±2.40 (14.5–20.80; 6 | 7.2 10.2 16.4 | 7.9 ±1.25 (6.2–9.8); 8 10.8 ±1.48 (9.0–13.6); 8 18.9 ±2.17 (16.2–21.8); 8 |
| Toe 6 Gonopore - Anus | ? 11.8 ±1.97 (9.1–15.1); 6 7.3 7.6 ±1.34 (5.8–9.6); 6 | 11.8 6.5 | 11.4 ±1.67 (9.6–14.6); 7 - |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |