Cyrtodactylus wayakonei
sp. n.
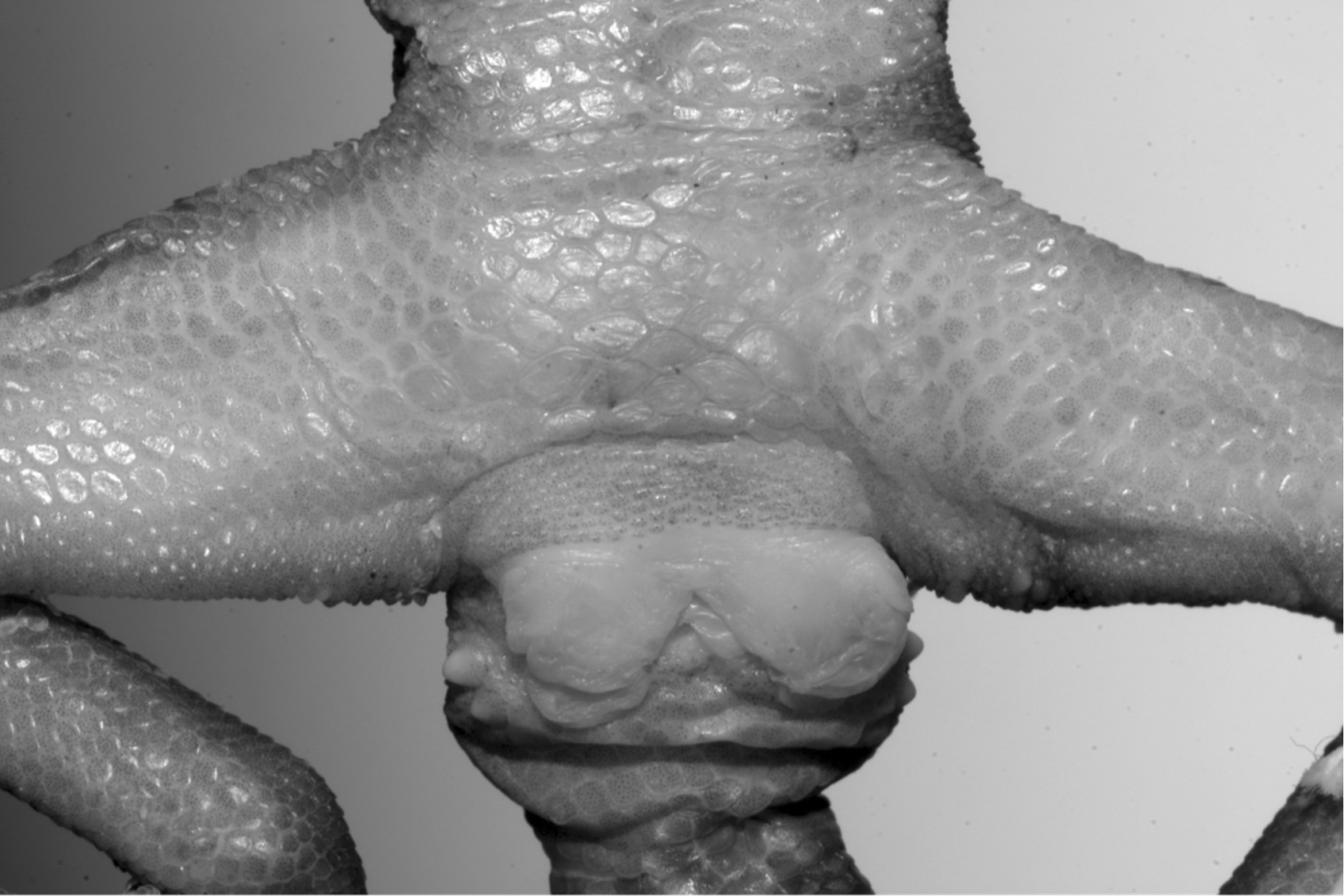
Holotype. IEBR A.2010.01, adult male from Kao Rao Cave, near Ban Nam Eng, Vieng Phoukha District, Luang Nam Tha Province, northern Laos (20o43.516’N, 101o09.239’E), altitude 730 m a.s.l., collected on 1 May 2010 by Truong Quang Nguyen and Phouthone Kingsada ( Figs. 1–3
View FIGURE 1
View FIGURE 2
View FIGURE 3
).
Paratypes. ZFMK 91016, adult male; MTD 47731, NUOL 2010.1, adult females; the same locality as the holotype, altitude 730–810 m a.s.l., collected between 1 and 2 May 2010 by Truong Quang Nguyen and Phouthone Kingsada ( Figs. 4–5
View FIGURE 4
View FIGURE 5
).
Diagnosis. A medium sized
Cyrtodactylus
of up to 90 mm maximum SVL, head distinct from slender body, with flat belly.
Cyrtodactylus wayakonei
sp. n. can be distinguished from all congeners on the basis of the following combination of characters: 7–8 supralabials; 9–10 infralabials; rostrum bearing smooth scales; 34–45 scales between fifth supralabials; dorsal tubercles smooth to slightly keeled; 17–19 rows of dorsal tubercles; ventrolateral folds slightly developed, with large tubercles; 31–35 ventrals; 85–98 scales around midbody; 151–163 scales between mental and cloacal slit; 6–8 precloacal pores in both sexes; precloacal groove lacking; femoral scales not enlarged; no femoral pores; subcaudals somewhat enlarged, broadened; tail tubercles flat, smooth; head and dorsum with blotched to reticulated pattern.
Description of holotype. Adult male with total length of 174.3 mm. Rostral wider than high, smaller than mental, dorsal margin straight, and medially with a straight, vertical rostral suture; 8 supralabials; 4 scale rows between supralabials and orbit; nares in contact with rostral, first supralabial on each side, and 4 nasal scales posteriorly; internasals lacking; snout bearing elongate medial impression; 45 scales between fifth supralabials; pupil vertical; anterior ciliaries 2–3 times as large as posterior ciliaries, without spinous tubercles; ear opening oblique, oval; interorbital region with small roundish, flat granular scales; larger tubercles dispersed within orbital region; granular scales on occiput; granular scales half the size of medial snout scales; head tubercles in temporal region twice the size of adjoining scales; scattered small tubercles in parietal region; few tubercles below a line connecting corners of mouth and ear opening; mental triangular, approximately as broad as long; 10 (right), 9 (left) infralabials; two postmentals, trapezoidal, approximately twice as long as broad, anteriorly in contact with mental and first infralabial; eight scales in contact with postmentals (except labials), outer ones distinctly larger, the same size of postmentals; gular scales granular.
Dorsal scales flat, arranged side by side, as large as medial snout scales, irregularly arranged in vertebral region, and laterally in relatively regular tranversal rows; dorsal tubercles medially oval, laterally roundish, blunt-conical, in part slightly keeled; dorsal tubercles 3–4 times as large as adjoining dorsal scales, arranged in 19 more or less regular longitudinal rows at midbody; dorsal tubercles surrounded by 9–10 dorsal scales, longitudinally separated by 2–3 and transversally separated by 2–4 dorsals; ventrolateral folds slightly developed, interspersed with large, roundish, domed to conical tubercles; tubercles separated from each other by 1–3 scales; ventral scales flat, more or less juxtaposed, as large as lateral tubercles (somewhat larger in abdomen); 31 ventrals between ventrolateral folds; 85 scales around midbody; 159 scales between mental and cloacal slit; scales on upper arm flat, smooth, anteriorly twice the size of posteriorly; forearm dorsally granular, with conical tubercles, 1.5–2 times the size of adjoining granular scales; femoral scales flat anteriorly, smooth, more or less juxtaposed, dorsally granular, with roundish, conical tubercles; no distinctly enlarged femorals; shank dorsally granular, with conical tubercles; fingers and toes free of webbing; claw bordered by two scales; finger I with 13 (right), 12 (left), finger II with 15, finger III with 17, finger IV with 18 (right), 17 (left), and finger V with 19 subdigital scales, of which finger I has 4 basally broadened lamellae, finger II 5, finger III 5, finger IV 6 (right), 5 (left) and finger V 6; toe I with 12 (right), 13 (left), toe II with 16, toe III with 18, toe IV with 20 (right), 19 (left), toe V with 18 subdigital scales, of which toe I has 4 (right), 5 (left) basally broadened lamellae, toe II 5, toe III 5 (right), 6 (left), toe IV 7, and toe V 5; the length formula of fingers is I <II <III <IV> V and toes I <II <III <IV> V; eight precloacal pores in a wide angular series; pores slit-like, arranged at the posterior margin of scales; three rows of enlarged scales present in posterior region of precloacal pores; 2/2 conical, thickened and obliquely arranged postcloacal tubercles. Original tail, in particular at base, with distinct whorls, medially with 5–6 dorsal scale rows, and with several tubercles in a single transversal row at the whorl’s end; tubercles flat, smooth, 1.5 times the size of lateral tubercles; dorsal scales flat, juxtaposed, of different sizes; lateral tail scales approximately same size; subcaudals flat, smooth; median rows of subcaudals for large parts somewhat enlarged.
Color in preservative (70% ethanol): Dorsum grey-brown; head surface with large, roundish, oval or somewhat asymmetrical blotches, with light margins; nasofrenal and postocular stripes lacking; a continuous, U-shaped nuchal band lacking as well; dorsum with reticulated grey-brown pattern and small, white blotches; extremities with small, white dorsal blotches; upper extremities with short, thin and light bands. Throat, belly and ventral side of extremities whitish-grey; dorsum of tail dark brown with six light transversal bands (with dark centre that fades towards the tail tip); tail venter dark grey-brown with thin light transversal markings corresponding to the light transversal bands of the dorsum.
The coloration in life is grey brown with dark purplish-brown blotches and banded to reticulated pattern; tubercles are beige to yellow as are the borders of the blotches on the head; the dorsum of the tail is greyishblack with beige pattern.
Variation. The paratype series largely corresponded with the description of the holotype. For measurements, scalation, and colour pattern variation see Figs. 4–5
View FIGURE 4
View FIGURE 5
and Table 1
View TABLE 1
. Characteristic are the blotches on the dorsal head surface with more or less bright light margins and the reticulated back pattern. Regenerated tails are dark, without pattern. The postcloacal tubercles of the males are somewhat larger than those of the females.
Further specimens. In March 2008, a male specimen of 90 mm snout-vent length was captured in Tham Phasat, northwest of Vieng Phoukha District, Luang Nam Tha Province, northern Laos. In this specimen, which largely agreed in colour pattern and morphology with the type series, the hemipenes could be everted ( Fig. 6
View FIGURE 6 a
). The fully everted hemipenes are slender, apically thickened; apex deeply bifurcated, labrum lacking; sulcus spermaticus thin, deep, bordered by bulging lips; sulcus branches broad, flat and stretching below the lobes towards the asulcal side, where they end; lobes without medial depression, with deep, partly elongate calyces with slightly denticulated margins.
Comparisons. The presence and arrangement of cloacal pores are the most important distinguishing characters in geckos. In the genus
Cyrtodactylus
, three different types occur: 1) different scales containing one pore each arranged in a more or less angular series anterior to the cloaca (= precloacal pores); 2) different scales containing one pore each, arranged in one or two series under the thighs and separated by poreless scales from the precloacal pores (= femoral pores); 3) precloacal pores and femoral pores forming a continuous, angular series (= precloacal-femoral pores) (see Rösler 2005). There are only few exceptions in which two of the aforementioned types occur together (for example
C. marmoratus
, see Rösler et al. 2007).
Cyrtodactylus wayakonei
sp. n. only has precloacal pores. Due to the lack of femoral pores it differs from
C. aaroni Günther & Rösler, 2003
(which has 9–12 precloacal pores + 8–13 femoral pores),
C. agusanensis (Taylor, 1915)
(8–11 + 8–11),
C. aequalis Bauer, 2003
(9 + 3–4),
C. auribalteatus Sumontha, Panitvong & Deein, 2010
(6 + 8–10),
C. baluensis (Mocquard, 1890)
(9–11 + 4–9),
C. biordinis Brown & McCoy, 1980
(11–14 + 15–28),
C. brevipalmatus (Smith, 1923)
(9–10 + 6–7),
C. capreoloides Rösler, Richards & Günther, 2007
(13 + 16–17),
C. erythrops Bauer, Kunya, Sumontha, Niyomwan, Panitvong, Pauwels, Chanhome & Kunya, 2009
(9 + 18–20),
C. gubernatoris (Annandale, 1913)
(9 + 6),
C. huynhi Ngo & Bauer, 2008
(7–9 + 1– 4),
C. louisiadensis ( De Vis, 1892)
(18–20 + 21–24),
C. mimikanus (Boulenger, 1914)
(7–17 + 7–18),
C. roesleri Ziegler, Nazarov, Orlov, Nguyen, Vu, Dang, Dinh & Schmitz, 2010
(20–28 in total),
C. russelli Bauer, 2003
(15 + 16–19),
C. salomonensis Rösler, Richards & Günther, 2007
(15 + 18–21),
C. slowinskii Bauer, 2002
(9–11 + 12),
C. takouensis Ngo & Bauer, 2008
(3–4 + 1),
C. tripartitus Kraus, 2008
(21–22 + 21–29),
C. wetariensis (Dunn, 1927)
(11 + 12–13) and
C. ziegleri Nazarov, Orlov, Nguyen & Ho, 2008
(5–8 + 1–3).
The following
Cyrtodactylus
species have precloacal-femoral pores, which are lacking in
Cyrtodactylus wayakonei
sp. n.:
C. batucolus Grismer, Chan, Grismer, Wood & Belabut, 2008
(having 43–46 precloacalfemoral pores),
C. chanhomeae Bauer, Sumontha & Pauwels, 2003
(32–34),
C. derongo Brown & Parker, 1973
,
C. deveti (Brongersma, 1948)
(18–23),
C. feae (Boulenger, 1893)
(32),
C. epiroticus Kraus, 2008
(60– 82),
C. halmahericus (Mertens, 1929)
(42–52),
C. jarujini Ulber, 1993
(52–54),
C. klugei Kraus, 2008
(66– 76),
C. loriae (Boulenger, 1898)
(30–81),
C. macrotuberculatus Grismer & Norhayati, 2008
(35–37),
C. novaeguineae (Schlegel, 1837)
(24–34),
C. phongnhakebangensis Ziegler, Rösler, Herrmann & Vu, 2003
(32– 42),
C. robustus Kraus, 2008
(75–85),
C. seribuatensis Youmans & Grismer, 2006
(40–44),
C. serratus Kraus, 2007
(87),
C. tiomanensis Das & Lim, 2000
(19),
C. variegatus (Blyth, 1859)
(32), and
C. zugi Oliver, Tjarhan, Mumpuni, Krey & Richards, 2008
(29–34).
The presence of precloacal pores in males distinguishes
Cyrtodactylus wayakonei
sp. n. from
C. gordongekkoi
( Das, 1993; see also Biswas, 2007), and
C. sermowaiensis
(de Rooij, 1915);
C. mandalayensis Mahony, 2009
has only 5 precloacal pores in a continuous wide angular series (versus 6–8 in the new species), plus a single additional enlarged pored scale bordering posteriorly apex of series angle.
Cyrtodactylus wayakonei
sp. n. lacks a precloacal depression, which completely or in part hides the precloacal pores; it thus differs from the following species, which have a precloacal depression:
C. aurensis Grismer, 2005
,
C. cavernicolus Inger & King, 1961
,
C. fumosus (Müller, 1895)
,
C. nuaulu Oliver, Edgar, Mumpuni, Iskandar & Lilley, 2009
,
C. marmoratus Gray, 1831
,
C. papuensis ( Brongersma, 1934)
,
C. philippinicus (Steindachner, 1867)
,
C. pubisulcus Inger, 1957
,
C. pulchellus Gray, 1828
,
C. rubidus (Blyth, 1860)
,
C. sadleiri Wells & Wellington, 1985
,
C. semenanjungensis Grismer & Leong, 2005
,
C. spinosus Linkem, McGuire, Hayden, Setiadi, Bickford & Brown, 2008
and
C. stresemanni Rösler & Glaw, 2008
.
Due to the presence of large tubercles on the lateral folds,
Cyrtodactylus wayakonei
sp. n. differs from
C. annandalei Bauer, 2003
,
C. ayeyarwadyensis Bauer, 2003
,
C. buchardi David, Teynié & Ohler, 2004
,
C. chrysopylos Bauer, 2003
,
C. consobrinoides (Annandale, 1905)
,
C. cracens Batuwita & Bahir, 2005
,
C. cryptus Heidrich, Rösler, Vu, Böhme & Ziegler, 2007
,
C. edwardtaylori Batuwita & Bahir, 2005
,
C. fraenatus (Günther, 1864)
,
C. gansi Bauer, 2003
,
C. interdigitalis Ulber, 1993
,
C. jellesmae (Boulenger, 1897)
,
C. laevigatus (Darevsky, 1964)
,
C. malcolmsmithi (Constable, 1949)
,
C. oldhami (Theobald, 1876)
,
C. peguensis (Boulenger, 1893)
,
C. quadrivirgatus Taylor, 1962
,
C. redimiculus King, 1962
,
C. soba Batuwita & Bahir, 2005
,
C. subsolanus Batuwita & Bahir, 2005
,
C. sumonthai Bauer, Pauwels & Chanhome, 2002
,
C. tautbatorum Welton, Siler, Diesmos & Brown, 2009
(46–58),
C. thirakhupti Pauwels, Bauer, Sumontha & Chanhome 2004
,
C. tigroides Bauer, Sumontha & Pauwels, 2003
,
C. tuberculatus (Lucas & Frost, 1900)
and
C. wakeorum Bauer, 2003
.
The size and thus the number of midbody ventrals in
Cyrtodactylus
varies between 22–82 (
C. variegatus
versus
C. caovansungi
, see Rösler & Glaw 2008).
Cyrtodactylus wayakonei
sp. n. belongs to the species with relatively large ventrals (<36) and thus differs from the following species, which have distinctly smaller and thus more ventrals:
C. adleri Das, 1997
(which has 48–50 ventrals),
C. agamensis (Bleeker, 1860)
(67),
C. annulatus (Taylor, 1915)
(50–60),
C. caovansungi Orlov, Nguyen, Nazarov, Ananjeva & Nguyen, 2007
(79– 82),
C. consobrinus (Peters, 1871)
(65–70),
C. eisenmanae Ngo, 2008
(44–45),
C. jambangan Welton, Siler, Diesmos & Brown, 2010
(48–63),
C. jarakensis Grismer, Chan, Grismer, Wood & Belabut, 2008
(61),
C. lateralis (Werner, 1896)
(60–64),
C. malayanus ( De Rooij, 1915)
(58–62),
C. matsuii Hikida, 1990
(51),
C. pantiensis Grismer, Chan, Grismer, Wood & Belabut, 2008
(40–46),
C. wallacei Hayden, Brown, Gillespie, Setiadi, Linkem, Iskandar, Umilaela, Bickford, Riyanto, Mumpuni & McGuire, 2008
(45–49), and
C. yoshii Hikida, 1990
(50–58).
Cyrtodactylus wayakonei
sp. n. differs from
C. angularis ( Smith, 1921)
,
C. intermedius (Smith, 1917)
,
C. cattienensis Geissler, Nazarov, Orlov, Böhme, Phung, Nguyen & Ziegler, 2009
,
C. hontreensis Ngo, Grismer & Grismer, 2008
, and
C. phuquocensis Ngo, Grismer & Grismer, 2010
by lacking enlarged femoral scales, and lacking dark band across the nape; in contrast to the new species,
C. leegrismeri Chan & Ahmad, 2010
,
C. sworderi (Smith, 1925)
,
C. yangbayensis Ngo & Chan, 2010
, and
C. zhaoermii Shi & Zhao, 2010
have a series of enlarged femoral scales.
Cyrtodactylus wayakonei
sp. n. has a lower number of enlarged dorsal tubercles (17–19) compared with
C. brevidactylus Bauer, 2002
(27), and
C. irianjayaensis Rösler, 2000
.
Cyrtodactylus wayakonei
sp. n. differs from
C. khasiensis (Jerdon, 1870)
by fewer precloacal pores (6–8 versus 12–14);
C. tamaiensis ( Smith, 1940)
in contrast has 40 precloacal-femoral pores ( Smith 1940, Mahony 2009).
Cyrtodactylus badenensis Nguyen, Orlov & Darevsky, 2006
differs from the new species by lacking precloacal pores, having a lower SVL (74 versus 90 mm) and a lower ventral count (25–28 versus 31–35) ( Nguyen et al. 2006).
Cyrtodactylus chauquangensis Hoang, Orlov, Ananjeva, Johns, Hoang & Dau, 2007
differs from
C. wayakonei
sp. n. by having more scales around the midbody (128–130 versus 85–98) ( Hoang et al. 2007).
Cyrtodactylus condorensis ( Smith, 1921)
has keeled snout scales (which are smooth in
C. wayakonei
sp. n.), and differs by a higher ventral count (35–40 versus 31–35) and fewer precloacal pores (4–7 versus 6–8) ( Smith 1921).
Cyrtodactylus darmandvillei ( Weber, 1890)
has dorsal tubercles, which are distinctly ribbed or uni- to three-carinate (versus smooth to indistinctly keeled dorsal tubercles in
C. wayakonei
sp. n.) and it differs from the new species by lacking precloacal pores, by having a lower SVL (75 versus 86.6 mm), and a higher ventral count (36–40 versus 31–35) ( Weber 1890, De Rooij 1915, Brongersma 1934).
Cyrtodactylus elok Dring, 1979
has more ventrals (44 versus 31–35), and spine-shaped tail tubercles, which are lacking in the new species (Dring 1979).
Cyrtodactylus grismeri Ngo, 2008
differs from
C. wayakonei
sp. n. by the absence of precloacal pores (0 versus 6–8), a higher supralabial count (10–13 versus 7–8) and a different dorsal pattern (the reticulated head pattern is lacking in
C. grismeri
and the tubercles of the vertebral region are pale) (Ngo 2008).
Cyrtodactylus ingeri Hikida, 1990
differs from
C. wayakonei
sp. n. by its smaller SVL (80.2 versus 90 mm) and a higher ventral count (40–43 versus 31–35). The dorsal pattern of
C. ingeri
is variable, with the postocular streak reaching in part beyond the thighs (e.g., Hikida 1990, Das 2004).
Cyrtodactylus irregularis ( Smith, 1921)
is distinguishable from the new species by a higher ventral count (38–46 versus 31–35), by having fewer precloacal pores (5–7 versus 6–8) and by the dorsal pattern, which consists of an U-shaped, undivided nuchal collar and large, asymmetrical dark blotches and bands in the former species, respectively ( Smith 1935, Nazarov et al. 2008).
Cyrtodactylus nigriocularis Nguyen, Orlov & Darevsky, 2006
differs from
Cyrtodactylus wayakonei
sp. n. by its higher ventral count (42–49 versus 31–35), by its lower number of precloacal pores (0–2 versus 6–8) and by its nearly patternless dorsum ( Nguyen et al. 2006).
Cyrtodactylus papilionoides Ulber & Grossmann, 1991
differs from
C. wayakonei
sp. n. by fewer precloacal pores (4–6 versus 6–8) as well as by its characteristic dorsal pattern, which consists of serrated broad dark bands with light margins or transversal rows of blotches ( Ulber & Grossmann 1991).
Cyrtodactylus paradoxus ( Darevsky & Szczerbak, 1997)
differs from the new species by the total absence of or the presence of only four precloacal pores ( Darevsky & Szczerbak 1997, Nguyen et al. 2006, Nazarov et al. 2008). In addition,
C. paradoxus
differs from
C. wayakonei
sp. n. by fewer scales between mental and cloacal slit (134–148 versus 151–163).
Cyrtodactylus pseudoquadrivirgatus Rösler, Vu, Nguyen, Ngo & Ziegler, 2008
differs from
C. wayakonei
sp. n. by having more scales between fifth supralabials (42–61 versus 34–45) and a higher ventral count (41– 57 versus 31–35) as well as by its dorsal pattern, which consists of continuous or interrupted stripes or bands in
C. pseudoquadrivirgatus (Rösler et al. 2008)
.
Cyrtodactylus cracens Batuwita & Bahir, 2005
,
C. edwardtaylori Batuwita & Bahir, 2005
,
C. fraenatus (Günther, 1864)
,
C. soba Batuwita & Bahir, 2005
and
C. subsolanus Batuwita & Bahir, 2005
have a well developed, non-denticulate lateral fold, which distinguishes them from
C. wayakonei
sp. n. ( Batuwita & Bahir 2005).
C. ramboda Batuwita & Bahir, 2005
differs from
C. wayakonei
sp. n., e.g., by a distinctly lower number of dorsal tubercles at midbody (9 versus 17–19) ( Batuwita & Bahir 2005).
Cyrtodactylus murua Kraus & Allison, 2006
differs from the new species by having widened subcaudals, a U-shaped nuchal band, and three dark dorsal bands with jagged margins that are medially partially split by a thin light stripe.
Cyrtodactylus wayakonei
sp. n. differs from the representatives of the subgenus
Geckoella
by a larger SVL (90 versus 52–80 mm) and by the presence of precloacal pores in both sexes from
C. (Geckoella) albofasciatus (Boulenger, 1885)
,
C. (Geckoella) deccanensis (Günther, 1864)
,
C. (Geckoella) jeyporensis (Beddome, 1877)
,
C. (Geckoella) nebulosus (Beddome, 1870)
,
C. (Geckoella) collegalensis (Beddome, 1870)
and
C. (Geckoella) yakhuna (Deraniyagala, 1945)
.
Cyrtodactylus (Geckoella) triedrus (Günther, 1864)
differs from the new species in having 3–4 precloacal and 3–4 femoral pores ( Das 1996, Rösler & Glaw 2008).
Systematics. Based on genital morphology, the genus
Cyrtodactylus
can be divided into two groups 1) members with comb-like calyces on the apex and in part also on the truncus, such as
C. phongnhakebangensis
(see Ziegler et al. 2002); 2) members which have apical calyces with lamellar skin bulges with denticulated margins, such as
C. pseudoquadrivirgatus (Rösler et al. 2008)
.
Cyrtodactylus wayakonei
sp. n. shows a normal development of calyces on the hemipenis and thus belongs to the first group.
The dorsal pattern of
Cyrtodactylus wayakonei
sp. n. most closely resembles
C. consobrinus
from West Malaysia, Borneo and Sumatra (see Welch 1994). However, the type series of
Cyrtodactylus wayakonei
sp. n. lacks pale banded extremities and has blotched arms and legs (see figures depicted in Manthey & Grossmann 1997, Chan-ard et al. 1999).
Cyrtodactylus consobrinus
from Borneo also shows banded extremities, but the species shows a broad variation in scalation such as in the presence/number of femoral scales ( Malkmus 1988, 1989, 1991, Hikida 1990).
C. consobrinus
is a scansorial forest dweller (e.g. Malkmus et al. 2002), whereas
C. wayakonei
sp. n. is cavernicolous and adapted to karst caves. Possible phylogenetic relationship of the aforementioned, superficially similar but geographically distant species have to be studied by additional data sets in the future, such as molecular approaches.
Another species which could to be close to
Cyrtodactylus wayakonei
sp. n. is
C. chauquangensis
, a hypothesis which also will be tested in future molecular phylogenetic approaches. Both species have a similar SVL, share the number of SPL, SBL, N, I, IO, PM, LZ1, LZ4, PP, have enlarged subcaudals under the tail, and do lack femoral pores and enlarged femoral scales. However,
Cyrtodactylus wayakonei
sp. n. differs from
C. chauquangensis
in the number of scales around the midbody and in mental shield and cloaca morphology ( Hoang et al. 2007, Nazarov et al. 2008). Amongst others,
C. wayakonei
sp. n. also has fewer ventrals than
C. chauquangensis
(31–35 versus 36–38).
C. chauquangensis
, which has a dorsal pattern consisting of three to four broad dark serrated bands with dark blotches in between, is a karst cave inhabitant as also is
C. wayakonei
sp. n. The type locality of
C. chauquangensis
is Chau Quang village, Quy Hop district, in Nghe An Province, northern Vietnam, about 430 km distant from the type locality of the new species. Further cavernicolous species were recently described from Vietnam, such as
C. huynhi
and
C. takouensis ( Ngo & Bauer 2008)
. Future molecular comparisons will show whether these “Indochinese” species with similar ecological adaptations are in fact phylogenetically close.
Etymology. The new
Cyrtodactylus
species is named after Associate Professor Dr. Sengdeuane Wayakone, Director, Office of Planning and International Cooperation, National University of Laos, in recognition of his efforts to investigate and protect biodiversity in Laos.
Distribution. The species currently is known only from few caves in Vieng Phoukha District, Luang Nam Tha Province, northern Laos ( Fig. 7
View FIGURE 7
).
Natural history. The type series was collected at night between 19–23:00 in the karst forest at elevations of 730–810 m ( Fig. 8
View FIGURE 8
). Except for the specimen NUOL 2010.1, which was collected on fallen tree, the other specimens were found in limestone crevices, about 1–1.5 m above the forest floor. In addition, two geckos were observed hanging on the cliff near the Kao Rao Cave ( Fig. 9
View FIGURE 9
). The female paratype NUOL 2010.1 contained two large, roundish to oval-shaped eggs of 10.5 mm maximum total length.
Of the specimens observed in March 2008, one adult male was seen at night at the entrance area of a limestone cave which is part of the Tham Phasat cave system in the middle part of a limestone hill covered with secondary evergreen forest (Phou Phasat). The male was sitting on a stone near the wall 1.5 m above the ground, 50 m away from the cave entrance. Another specimen seen in March 2008 was observed in the small back entrance of the big cave (Tham Phasat Thia) at the base of this hill. This animal was found on the same day at 9 am sitting in a narrow and deep hole at the ceiling of the cave 6 m away from the entrance. At 50 m inside the cave at 11 am, a temperature of 19°C was measured, outside 26°C. A third specimen seen in March 2008 was discovered on 5 March at 6 pm in a side entrance of the Tham Nam Eng cave that was surrounded by a small patch of dense secondary evergreen forest. The gecko was hiding in a small hole at the ceiling of the high entrance hall of this cave ca. 20 m away from entrance. We did not observe any
Cyrtodactylus
outside the caves although many crevices and holes were available in both localities.