
Isophya andreevae Peshev, 1981
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3658.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C02D1C74-25C0-41DD-B098-62098EB7B62A |
|
DOI |
https://doi.org/10.5281/zenodo.5617337 |
|
persistent identifier |
https://treatment.plazi.org/id/F26F3128-391D-FF8A-B1B0-0CE6FAC69981 |
|
treatment provided by |
Plazi |
|
scientific name |
Isophya andreevae Peshev, 1981 |
| status |
|
Isophya andreevae Peshev, 1981
( Figs 10, 11 View FIGURES 1 – 18 , 38, 63 View FIGURES 56 – 79 , 87 View FIGURES 80 – 104 , 112 View FIGURES 105 – 129 , 142, 143 View FIGURES 139 – 144 , 148, 149 View FIGURES 145 – 149 , 192 View FIGURE 192 )
Isophya andreevae Peshev : Peshev 1981 (sp.n.).
Morphological description: Peshev 1981. Karyotype: Warchałowska-Śliwa et al. 2008.
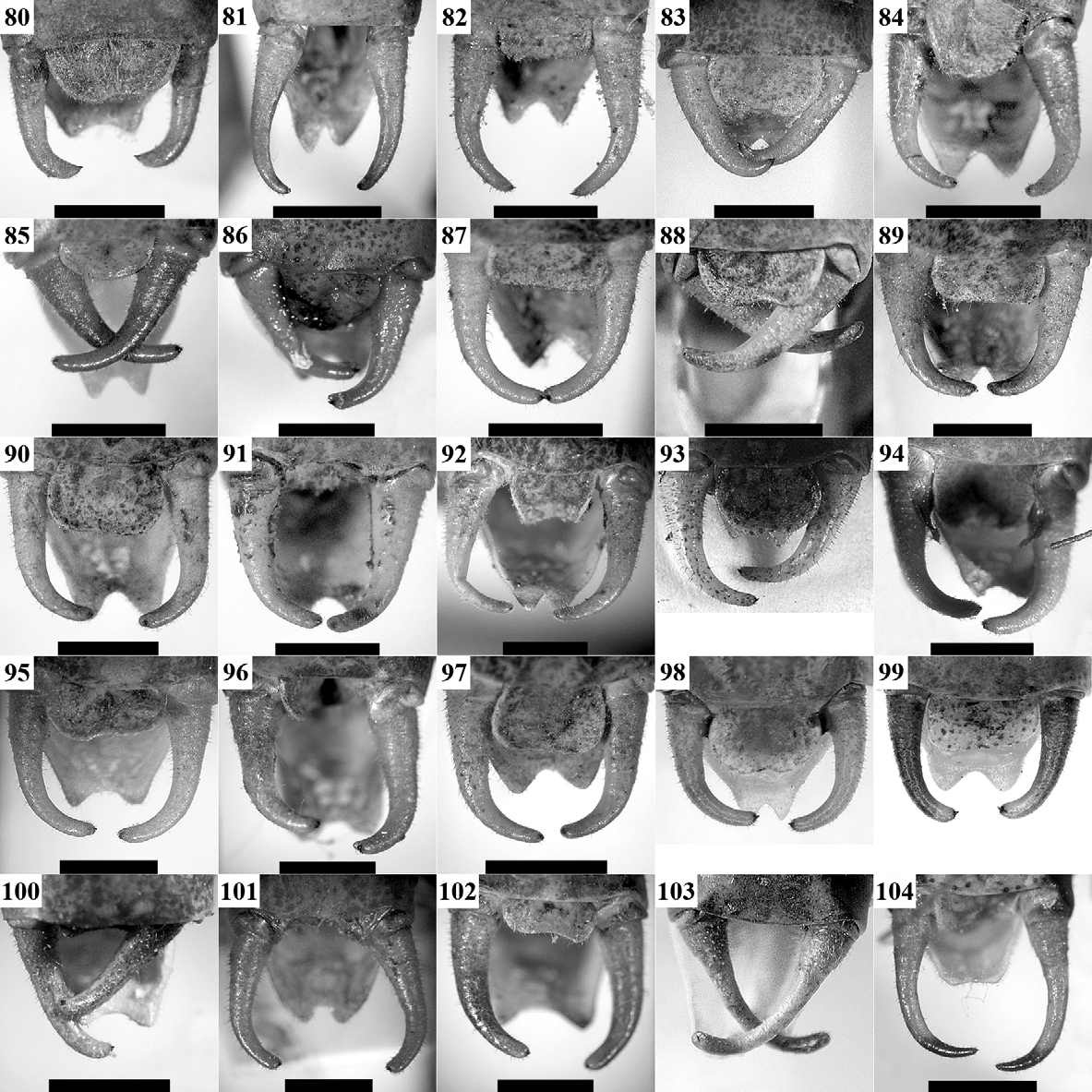
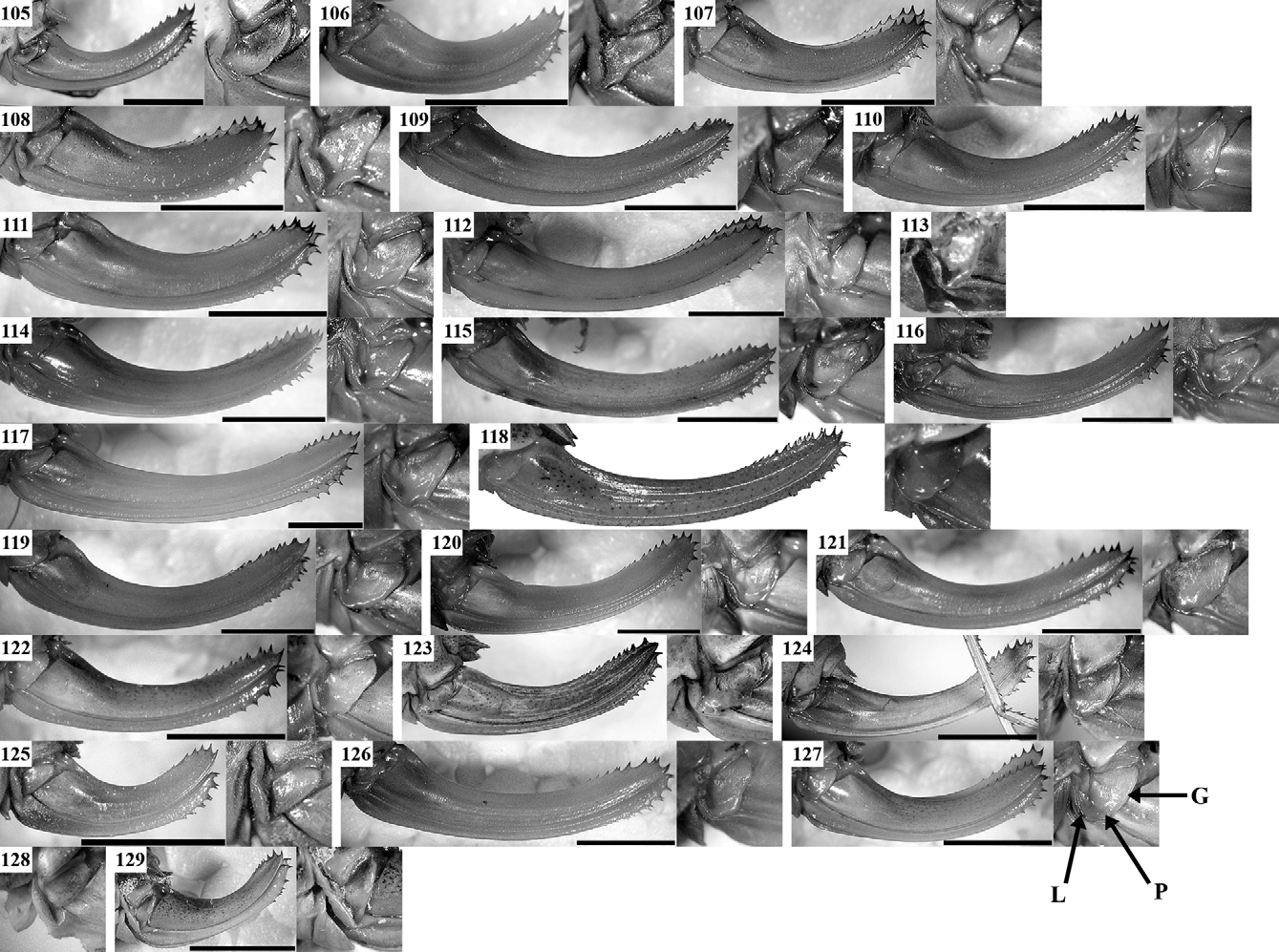
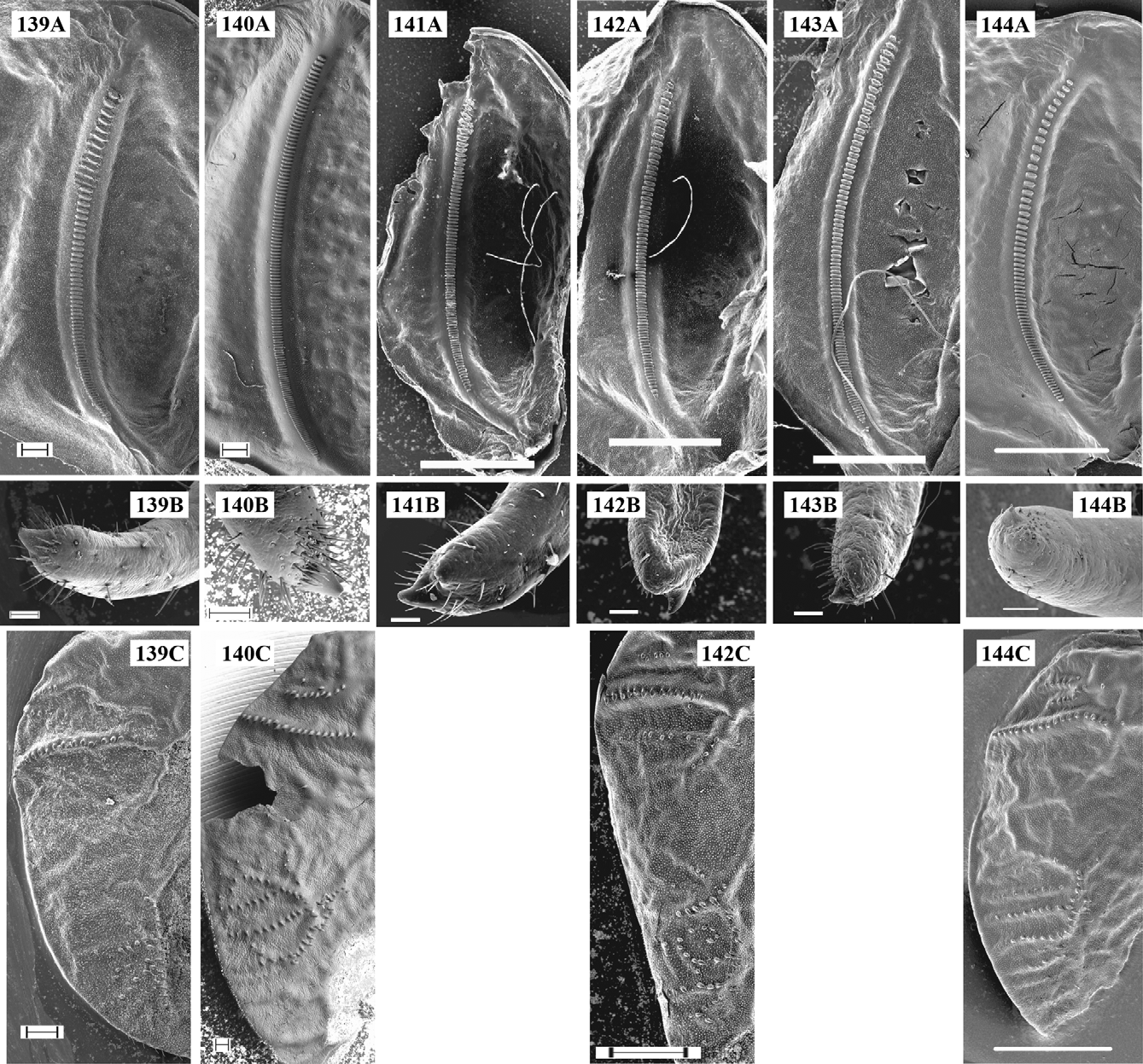
Supplement to the description and a diagnosis: Body colouration yellowish- or light-green. The tibiae, tarsi, and antennae may be yellowish. The disc of male tegmina is yellowish- or greenish-brown with lighter (usually yellowish) CuP. CuP is wide, strong. The stridulatory file ( Figs 142 View FIGURES 139 – 144 A, 143A) has a length of 2.9–3.6 mm with 77– 92 teeth in studied specimens. Female stridulatory apparatus is shown in Fig. 142 View FIGURES 139 – 144 C. The cercal tooth ( Fig. 142 View FIGURES 139 – 144 B, 143B) is long, stout, the excision between the tooth and cercus apex is less expressed than in I. tosevski .
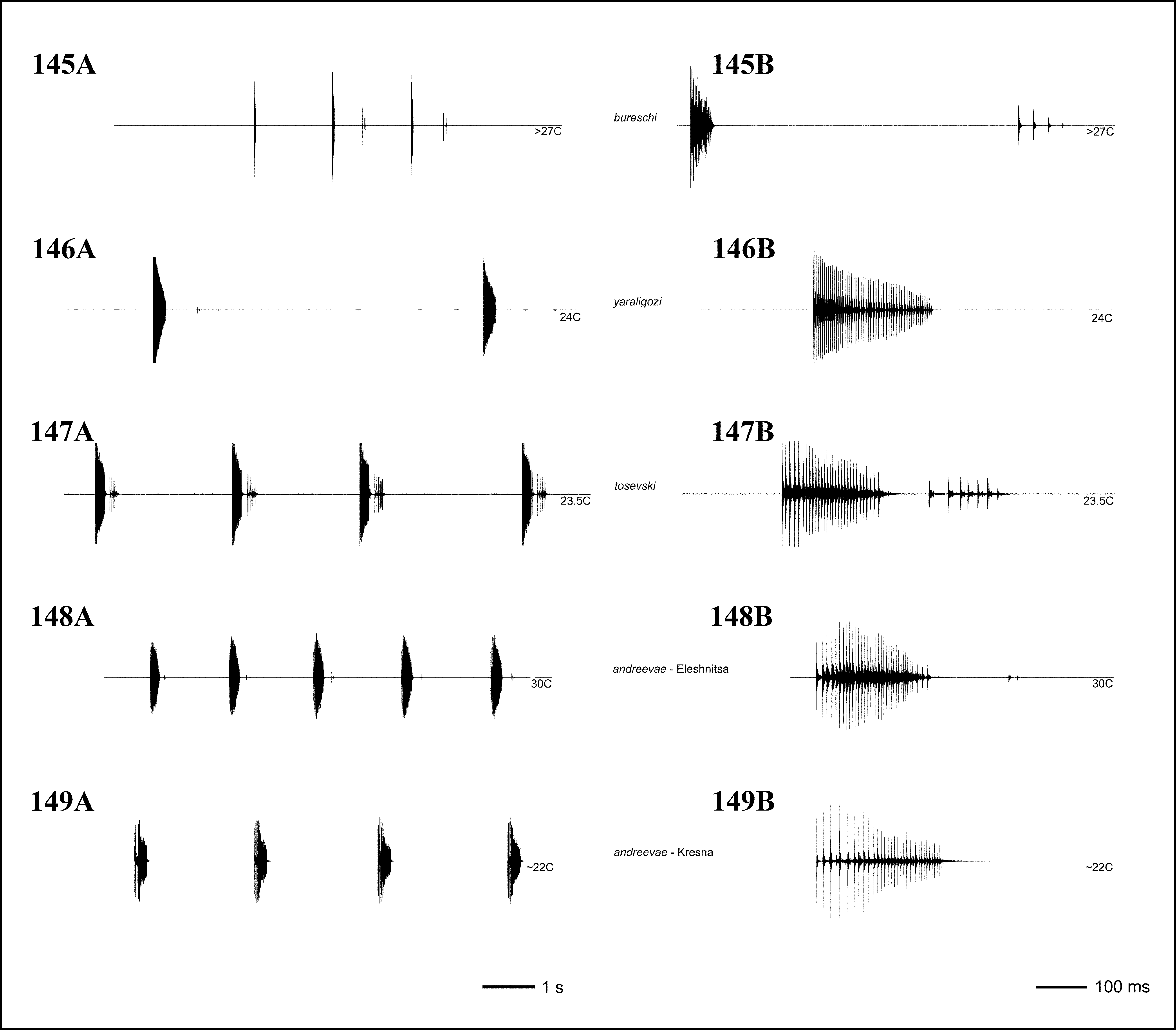
Bioacoustics: The studied populations (from the Strouma Valley: Kresna Gorge, and Rila Mountains: Eleshnitsa Locality) show a similar song including groups of (2–)3–6 (rarely single) syllables comprising well separated impulses. A tendency for occurrence of after-clicks was observed. Duration of the syllables’ and impulses’ period is influenced by body temperature, which may be the main reason for temporal differences between the songs of both populations. Heller (1988) showed oscillograms of two types of songs of I. tosevski . The oscillogram on Abbildung 30-C (p. 228) fits well the song of I. andreevae .
At 20–24°С the song of the lowland population (Strouma valley) ( Fig. 149 View FIGURES 145 – 149 ) had the following characteristics: the syllables within the groups are separated by intervals of 2.5– 3.5 s. The syllable was not split into two parts, lasted about 250–450 ms (mean 324±67) and consisted of 26–38 impulses (mean 33±4). First 8–12 impulse periods were considerably longer than the following; generally the impulse period strongly varied and had a mean value of 9.9 ms. The impulse period measured within six syllables in one individual at 20°С lasted 3–31 ms (mean 9.5±5.3; median=8; n=168). This pattern fits well with the oscillogram shown by Heller (1988) on Abbildung 30C (p. 228; specimen from Greece, Kilkis distr., Paikon Mt.). Though the shown syllable is shorter (about 200 ms) it is surely due to higher recording temperature (27°С). Therefore, there is a possibility that this recording belongs to I. andreevae .
At 30°С the mountain population (Rila Mts) ( Fig. 148 View FIGURES 145 – 149 ) showed shortened interval between syllables within the groups (lasting 1–2 s) and the interval between impulses within the syllable, though the tendency for longer impulse period in the beginning of the syllables remained. Here, a trend for splitting the syllables of a main part and after-clicks appeared, yet, the presence/absence of after-clicks vary within a syllable group (usually the last syllables of a group had better expressed additional impulses). It is not clear, however, whether the appearance of after-clicks depends on the body temperature, the age of specimens or/and is a population feature. The main part of the syllables lasted 180–250 ms (mean 218±15) and included 36–42 impulses (mean 40±2). The impulse period lasted 3–13 ms (mean 5.5). When after-clicks were present (1–5, usually 1–3 in number) the syllable had a length of 300–500 ms (mean 395±47). Thus, this population shows a song similar to the main syllable part of I. tosevski .
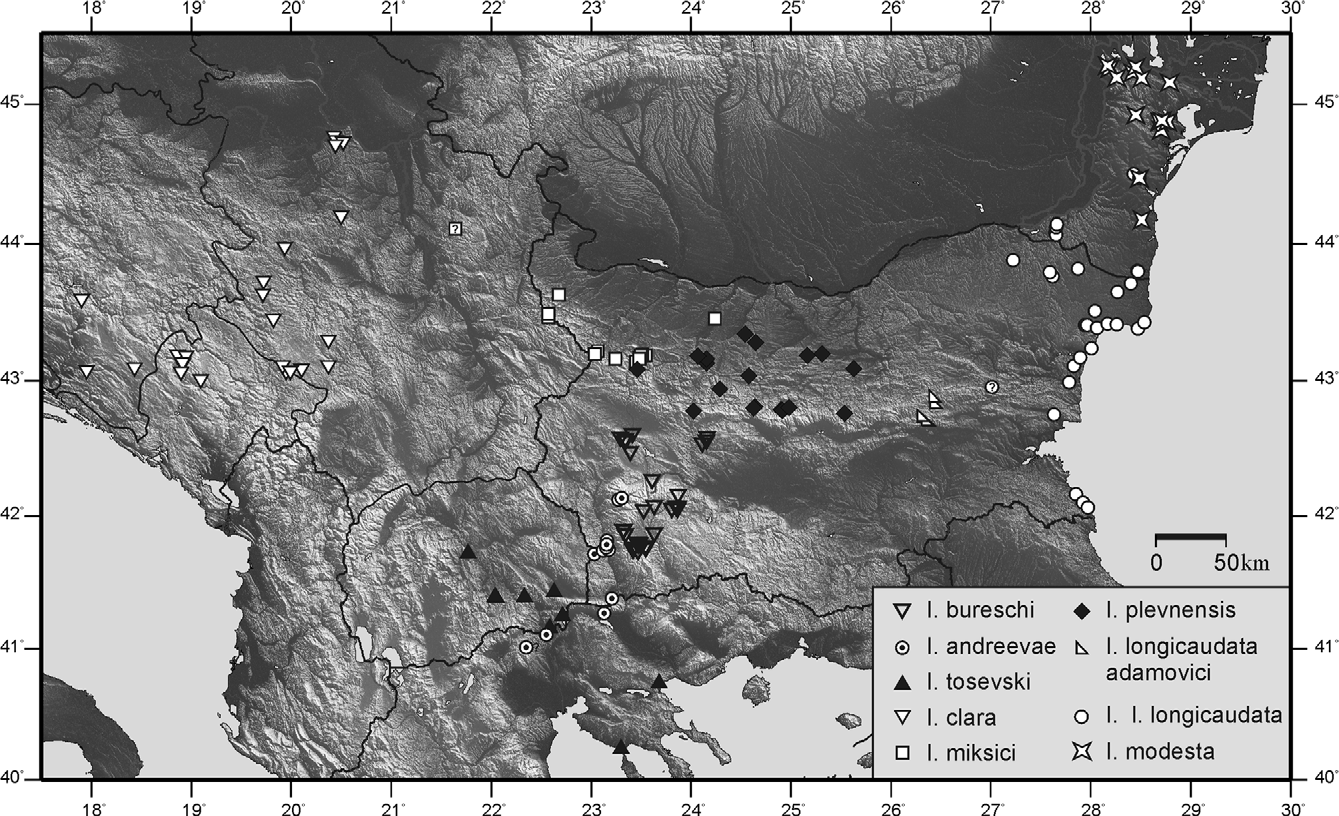
Distribution ( Fig. 192 View FIGURE 192 ) and phenology: This endemic species was found along the middle course of Strouma River and the neighbouring mountain slopes between 300 and 1800 m in Rila, Maleshevska Planina, Belassitsa (SW Bulgaria), and possibly Paikon Mountains in Greece. In the lowlands it appears as early species inhabiting mesophyte grass-shrub associations becoming xerophyte in the early summer, when the species disappear, while in the mountains it occurs in mesophyte meadows and forest clearings. Nymphs—III–V(–VI), imago—V–VII(–VIII). Notes to the literature distribution data: According to the oscillogram presented the record by Heller (1988) for I. tosevski from Paikon Mt. in Greece may be tentatively refered to I. andreevae (see above). However, the recorded specimen was studied (thanks are due to K.-G. Heller) and morphologically it cannot be distinguished from I. tosevski . Yet, as already stated, both species resemble each other very much and in this case we cannot judge whether the populations of these taxa occur sympatrically or even hybridise. Further, the case may concern either I. andreevae , or a hybrid, or the song of both taxa may show variations and thus cannot be distinguished.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |