
Grammatonotus bianchi, Lisher & Thein & Psomadakis, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4996.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:185BA4A5-3EFD-415E-BFD4-5E5279D83674 |
|
persistent identifier |
https://treatment.plazi.org/id/F423EC22-FFE5-FFB7-AEE1-7830F1B3FCBA |
|
treatment provided by |
Plazi |
|
scientific name |
Grammatonotus bianchi |
| status |
sp. nov. |
Grammatonotus bianchi sp. nov.
(English name: Bianchi’s splendid perch; FAO name: Andaman splendid perch; Burmese name: Andaman Yaung Sone Ka Ka Tit)
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Tables 1–3)
urn:lsid:zoobank.org:act:A9367510-D530-4E43-A3F6-5CB84702D066
Grammatonotus sp. : Psomadakis et al. 2019: 399, Pl. XXIX, fig. 219 ( Myanmar coast).
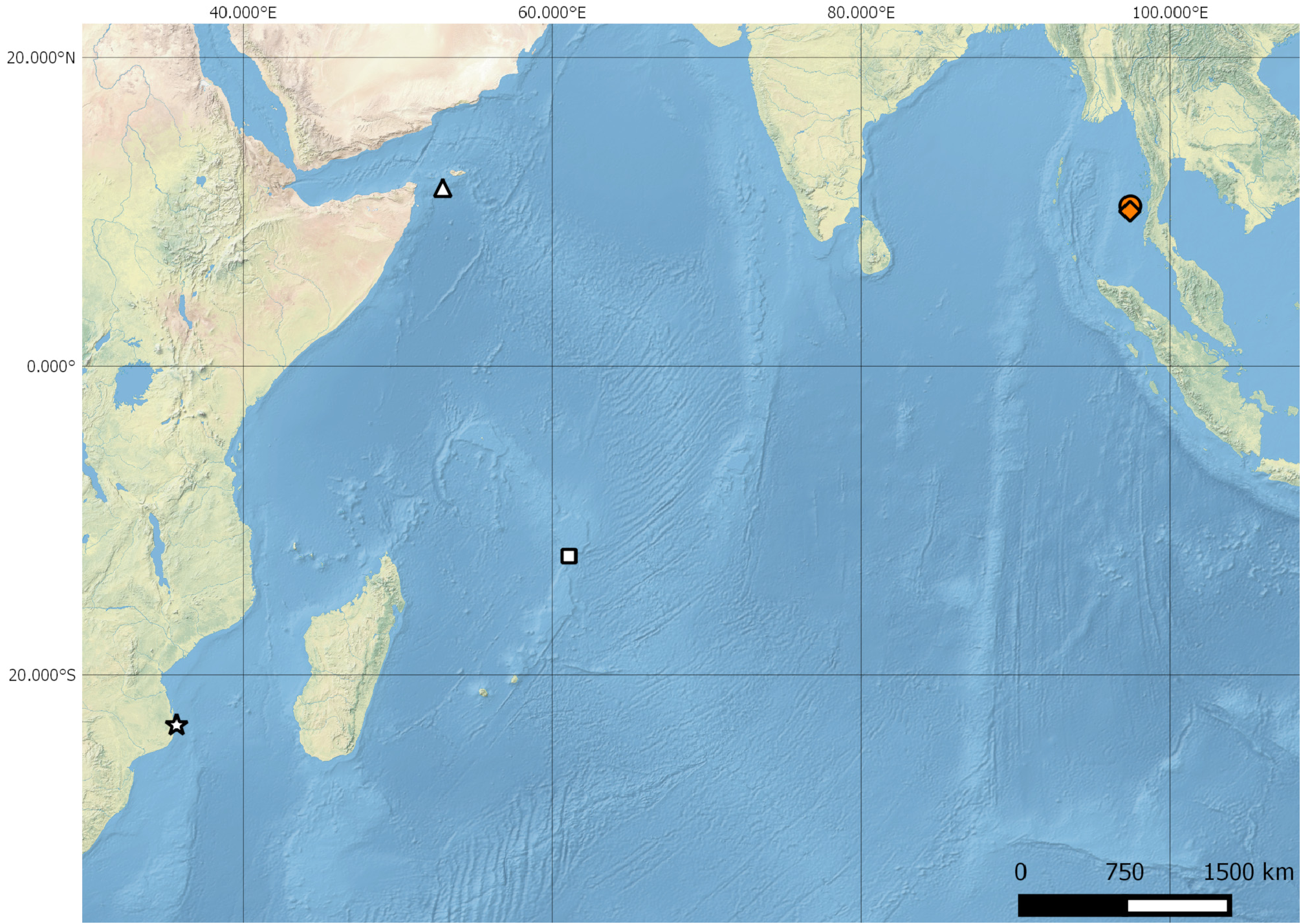
Holotype. SAIAB 208486 View Materials , 68.7 mm SL, off Tanintharyi coast, Myanmar, Andaman Sea , Indian Ocean, 10°2’42.00”N and 97°22’45.00”E, R / V Dr. Fridtjof Nansen, bottom trawl, station 154, 184 m depth, 27 September 2018, collected by P.N. Psomadakis. GoogleMaps
Paratype. SAIAB 208488 View Materials , 45.9 mm SL, same collection data as holotype GoogleMaps .
Photographic record (non-type). 121 mm TL, off Tanintharyi coast, Myanmar, Andaman Sea , Indian Ocean, 10°23’19.80”N and 97°24’48.60”E, R / V Dr. Fridtjof Nansen, bottom trawl, station 172, 182– 184 m depth, 28 May 2015, specimen lost GoogleMaps .
Diagnosis. A species of Grammatonotus distinguishable from its congeners by the following combination of characters: large head, its length 37.7–38.6% SL; postorbital head length 16.7–17% SL; large orbit, its diameter 14.4–15.3% SL; short caudal peduncle, its length 17.3–18.7% SL; caudal fin with upper and lower lobes produced; fresh coloration distinctive with unique bright yellow caudal fin covered by reddish spots and fuchsia or violet distal third (see Table 3, Fig. 2 View FIGURE 2 and detailed color description below).
Description. Morphometric and meristic data are presented in Table 1 and 2. Holotype data is presented first, followed by paratype data (if different) in parentheses. Body somewhat compressed, moderately deep, its depth 2.9–3.1 in SL; body width 5.9–6.3 in SL. Dorsal profile convex from snout tip to origin of first dorsal-fin spine, thereafter gradually convex to end of dorsal-fin base. Ventral profile concave from tip of lower jaw to pelvic-fin origin, thereafter parallel to body axis until anal-fin origin. Anal-fin base slightly concave. Caudal peduncle with slightly concave dorsal and ventral profiles. Head large, its length 2.6–2.7 in SL. Orbit large, its diameter 2.5–2.6 in HL. Mouth terminal and oblique; moderately large with an upper jaw length 2.5–2.6 in HL; jaws subequal. Maxilla reaching posteriorly to middle of eye. Premaxilla protrusile. No supramaxilla. Interorbital convex. One opercular spine; distal margins of interopercle and subopercle smooth; margin of preopercle smooth.
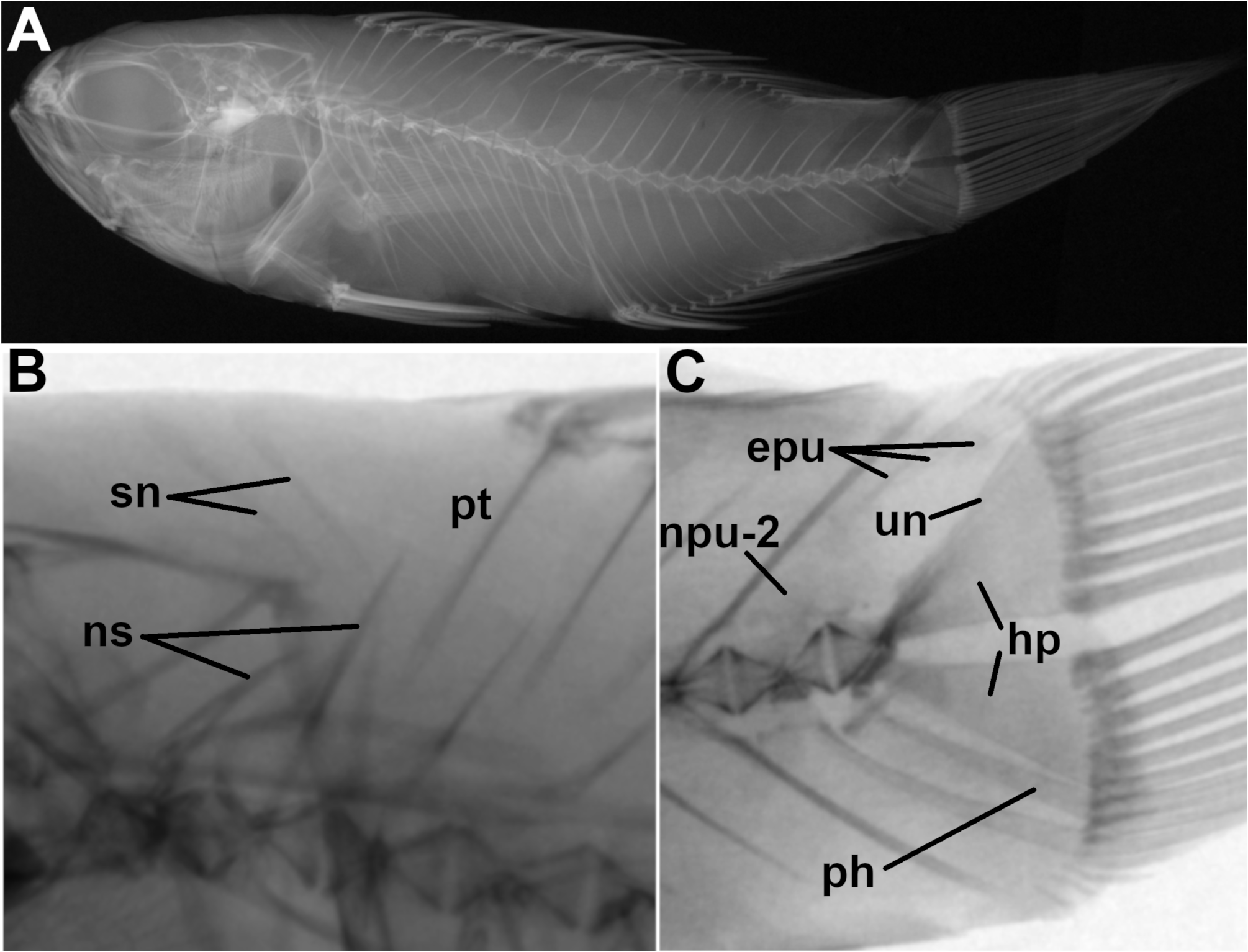
Dorsal fin not incised at junction of spinous and soft portions. Dorsal-fin rays XI, 9. Anal-fin rays III, 9. Pectoral-fin rays 20 (19). Pelvic-fin rays I, 5. Caudal fin rounded (somewhat pointed with median rays produced in lost specimen Fig. 2B View FIGURE 2 ) with filamentous rays on upper and lower lobes. Principal caudal-fin rays 15 (8+7); branched caudal-fin rays 13 (7+6); procurrent caudal-fin rays 6 dorsally and ventrally. Branchiostegal rays 6. Pseudobranchs with 14 (12) filaments. Gill rakers long and slender, 8 (7) + 18, total number on first gill arch 26 (25). Lateral line count not possible because most scales missing; nine tubed lateral line scales remaining on left side of holotype from below fourth dorsal-fin spine to second dorsal-fin ray; 9 tubed lateral line scales remaining on right side of paratype from below the seventh dorsal-fin spine to the fifth dorsal-fin ray where the lateral line terminates. Mid-lateral series scales ca. 24. Circum-caudal-peduncular scales ca.16.
Vertebrae 24 (10 precaudal + 14 caudal). Parapophyses absent from first caudal vertebra. Supraneurals 2. One trisegmental pterygiophore associated with dorsal fin, and one with anal fin. Pleural ribs 8 pairs on vertebrae 3 to 10. Hypural fusions 1 w/2; 3 w/4. Epurals 3. Epineurals at least 11 pairs.
Premaxilla with outer series of canine-like teeth with three to four slightly enlarged at anterior end of jaw; inner band of villiform to small conical teeth, band expanded near symphysis; no teeth at symphysis. Dentary with series of conical teeth substantially smaller in size (less than half) to those on the premaxilla; two enlarged into caniniform teeth about one third back on jaw; patch of villiform teeth next to symphysis; one exserted canine at anterior end of jaw; no teeth at symphysis. Vomer with few small conical teeth arranged in a chevron-shaped patch, patch without posterior prolongation. Palatine with band of villiform teeth. No teeth on tongue or pterygoids.
Scales peripheral ctenoid. Body with mid-lateral series of modified scales (see Anderson et al. 2018, Fig.1C– D View FIGURE 1 ). Most of head, including maxilla, dentary, dorsum of snout, and interorbital region with scales. Dorsal, anal, pectoral and pelvic fins without scales; modified scales (interpelvic process) overlapping pelvic-fin bases along midventral line.
Fresh coloration described from photos including the holotype ( Fig. 2A View FIGURE 2 ), paratype and photographic record of one specimen ( Fig. 2B View FIGURE 2 ) not retained. Dorsal part of body reddish orange, fading to yellowish or pink on the sides; lower part of head and body whitish anteriorly on breast, belly (partially) and opercular region, becoming yellowish or pale pink behind pelvic-fins bases to caudal-fin base; anterior tip of upper jaw yellowish with reddish premaxilla ventral edge, lower jaw anterior half reddish; iris of eye yellow centrally with red peripherally almost reaching pupil both dorsally and ventrally, bluish band over red dorsal part of iris; pectoral fin semi-translucent and uniformly pale orange; dorsal fin bright yellow with reddish spots basally on interspinous membranes, a centrally located row of reddish spots from the fourth dorsal-fin spine, and reddish tips on dorsal-fin spines; pelvic fin mostly translucent with outer rays fuchsia or violet distally; anal fin pinkish basally and bright yellow distally; caudal fin bright yellow with reddish spots, distal third fuchsia or violet, filamentous upper and lower lobe rays yellow.
Preserved coloration. Uniformly yellowish brown throughout, opercle semi-translucent.
Etymology. The species is named in honor of Dr. Gabriella Bianchi, in recognition of her remarkable contribution to advancing knowledge on the marine resources and ecosystems of developing countries throughout a long and productive career at FAO in the role of Senior Fisheries Officer and now as Research Coordinator of the EAF- Nansen Programme. Early in her career, Gabriella was a key person in the FAO FishFinder Programme (formerly SIDP) and contributed significantly to our knowledge of the world’s marine biodiversity through the production of reference publications, including the FAO Species Identification Sheets for the Western Indian Ocean and for the Eastern Central Atlantic, as well as several FAO field species identification guides covering the fishery resources of many countries in Africa and Asia. The specific epithet is treated as a noun in apposition.
Distribution. Grammatonotus bianchi sp. nov. is currently known only from the type locality, off Myanmar’s Tanintharyi coast, in the Andaman Sea ( Fig. 1 View FIGURE 1 ) at depths of 182– 184 m.
Comparisons. Grammatonotus bianchi sp. nov. is distinguishable from all valid species of Grammatonotus by having a larger head, its length 37.7–38.6% SL [vs. 28.3–37.5% SL for all other congeners]; a larger postorbital head length 16.7–17.0% SL [vs. 10.7–16.2% SL]; a larger orbit diameter 14.4–15.3% SL [vs. 7.5–14.9% SL, except for G. macrophthalmus ]; a shorter caudal peduncle length 17.3–18.7% SL [vs. 20.9–27% SL, except for G. ambiortus and G. macrophthalmus ]; a unique fresh coloration with a bright yellow caudal fin covered by reddish spots and fuchsia or violet distal third (see Fig. 2 View FIGURE 2 , Table 3 and color description), and a rounded (semi-produced central rays in unretained specimen) caudal fin with produced upper and lower lobes (except for G. laysanus ).
It can be further distinguished from the only other described species of Grammatonotus occurring in the Indian Ocean, G. lanceolatus , by having a larger interorbital width 6.3–8.0% SL [vs. 5.3% SL]; shorter mid-caudal-fin rays 29–33.6% SL [vs. 38.9–41.9% SL], and slightly longer anal-fin base, its length 19.2–19.8% SL [vs. 18.6% SL].
Grammatonotus bianchi sp. nov. can be easily further distinguished from the two putative undescribed species from the southwestern Indian Ocean, namely, Grammatonotus sp. 1 (Mascarenes) and Grammatonotus sp. 2 ( Mozambique) sensu Anderson et al. (2018) by having a deeper body, its depth 32.0–34.6% SL [vs. 28.1–32.7 and 29.8–30.3% SL]; a longer upper jaw, its length 14.8–15.3% SL [vs. 12.4–14.2 and 13.0–14.2]; longer analfin spines, first spine length 5.5–5.9% SL [vs. 2.4–4.4 and 3.9–4.4 % SL], second spine length 10–10.2% SL [vs. 5.1–7.1 and 7.5–7.9], and third spine length 12.1–13.5% SL [vs. 6.1–8.7 and 9.3–9.7% SL].
When compared to the three Coral Triangle Grammatonotus species , G. bianchi sp. nov. can be further separated from G. brianne by a longer predorsal-fin length, 36.4–38.1% SL [vs. 30.6–33.5% SL]; a longer upper jaw length, 14.8–15.3% SL [vs. 12.6–14.2% SL]; substantially shorter mid-caudal-fin rays, length of 29–33.6% SL [vs. 58.6–82.2% SL]; longer anal-fin spines, with first spine length 5.5–5.9% SL [vs. 2.8–3.4% SL], second spine length 10.0–10.2% SL [vs. 5.3–7.3], and third spine length of 12.1–13.5% SL [vs. 7.5–9.2% SL]. From G. crosnieri by having a longer snout, its length 5.7–6.1% SL [vs. 4.3–5.4% SL]; shorter anal-fin spines, with first spine length 5.5–5.9% SL [vs. 6.2–10.3% SL], second spine length 10–10.2% SL [vs. 9.4–14.3% SL], and third spine length of 12.1–13.5% SL [vs. 11.1–15.3% SL]. From G. roseus by having a shallower body, its depth 32–34.6% SL [vs. 35.4–37.2% SL]; a shorter pectoral fin, its length 24.6–24.9% SL [vs. 25.0–26.1% SL]; a longer pelvic fin, its length 27–27.4% SL [vs. 24.8–25.7% SL].
When compared to the open Pacific Ocean species, G. bianchi sp. nov. can be further distinguished from G. laysanus by having a longer predorsal-fin length, 36.4–38.1% SL [vs. 30.7–36.8% SL]. From G. macrophthalmus by a longer predorsal-fin length, 36.4–38.1% SL [vs. 30.5–35.9% SL]; a longer upper jaw, its length 14.8–15.3% SL [vs. 11.6–14.1% SL]; a longer pectoral fin, its length 24.6–24.9% SL [vs. 18.9–23.7% SL]; longer second and third anal-fin spines, their length 10–10.2 and 12.1–13.5% SL respectively [vs. 7.7–8.7 and 9.5–11.5% SL]. From G. pelipel by a larger interorbital width of 6.3–8% SL [vs. 5.9–6.8% SL]; a longer upper jaw, its length 14.8–15.3% SL [vs. 12.8% SL]; shorter pectoral and pelvic fins, their lengths 24.6–24.9 and 27–27.4% SL [vs. 27.4 and 29.6% SL]; a deeper caudal peduncle, its depth 14.7–15.5% SL [vs. 12.5–14.4% SL]; longer mid-caudal-fin ray length of 29–33.6% SL [vs. 26.3–28.5% SL]; longer third anal spine, its length 12.1–13.5% SL [vs. 10.3% SL]. From G. surugaensis by having a shallower body, its depth 32–34.6% SL [vs. 35.7% SL]; a longer predorsal-fin length of 36.4–38.1% SL [vs. 31% SL]; a longer length of upper jaw 14.8–15.3% SL [vs. 13.2% SL]; shallower caudal penduncle, its depth 14.7–15.5% SL [vs. 17.5% SL]; longer depressed anal-fin length of 35.7–38.3% SL [vs. 31.6% SL]; longer third anal-fin spine, its length 12.1–13.5% SL [vs. 7.8% SL]; and longer anal-fin base, its length 19.2–19.8% SL [vs. 15.2% SL]. From G. xanthostigma by a deeper body, its depth 32–34.6% SL [vs. 29.6–31.1% SL]; a longer predorsal length, 36.4–38.1% SL [vs. 33.2–33.9% SL]; a longer snout, its length of 5.7–6.1% SL [vs. 4.7–4.9% SL]; a larger interorbital width of 6.3–8% SL [vs. 5.6–5.7% SL]; a longer upper jaw, its length 14.8–15.3% SL [vs. 12.3– 12.8% SL]; a deeper caudal peduncle, its depth 14.7–15.5% SL [vs. 12.6–13.8% SL]; substantially shorter mid-caudal-fin rays, their length 29–33.6% SL [vs. 55.6–59.3% SL]; shorter depressed anal fin, its length 35.7–38.3% SL [vs. 39.7–42.7% SL]; longer anal-fin spines, first spine length 5.5–5.9% SL [vs. 3.7–3.8% SL], second spine length 10–10.2% SL [vs. 6.7–7.2% SL], and third spine length of 12.1–13.5% SL [vs. 8.0–8.7% SL].

| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Grammatonotus bianchi
| Lisher, Mark W., Thein, Htun & Psomadakis, Peter N. 2021 |
Grammatonotus sp.
| Psomadakis, P. N. & Thein, H. & Russell, B. C. & Tun, M. T. 2019: 399 |