
Xenobatrachus lanthanites, Günther, Rainer & Knop, Ronny, 2006
|
publication ID |
https://doi.org/10.5281/zenodo.173194 |
|
DOI |
https://doi.org/10.5281/zenodo.5631776 |
|
persistent identifier |
https://treatment.plazi.org/id/F51187F8-7154-FF86-B17A-16CFFCC14C7B |
|
treatment provided by |
Plazi |
|
scientific name |
Xenobatrachus lanthanites |
| status |
sp. nov. |
Xenobatrachus lanthanites View in CoL sp. nov. ( Figs 3–10 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Holotype: ZMB 69558 (field number = FN 7788); adult male ( Figs 3 View FIGURE 3 and 4 View FIGURE 4 ), collected by R. Günther and H. Hasser on 9 July 2003 near the road to Ambaiduru Village, about 17 km direct line NE of Serui, Amoman Mountain, 1°45´S and 136°19´E, altitude 1100 m a.s.l., Yapen Island, Papua Province, Indonesia.
Paratypes: ZMB 69557 (FN 7787), ZMB 69559 (FN 7789), ZMB 69560 (FN 7790) and ZMB 69561 (FN 7791). All paratypes were collected near the locus typicus at the same day and from the same collectors as for the holotype. All paratypes are adult males; ZMB 69561 was cleared and doublestained as a bonecartilage preparation.
Diagnosis. With an average snouturostyle length of 21.9 mm the new species is clearly smaller than almost all other Xenobatrachus species, of which the smallest exhibit an average length of at least 27 mm. The only exception is Xenobatrachus anorbis Blum & Menzies which has about the same SVL (21.3–23.4, mean 22.5 mm) as Xb. lanthanites sp. nov. However, Xb. anorbis has conspicuously shorter tibiae than the new species (TL/SUL 0.29–0.32 vs. 0.40–0.42), a different ratio END / IND (1.26–1.32 vs. 0.94–1.20) and lacks digital discs completely (all data on Xb. anorbis according to Blum & Menzies 1989). Advertisement calls of the new species are long series of short piping notes, average note length is 121 milliseconds with a dominant frequency of about 1.0 kHz, note repetition rate is 1–2 notes per second. In external morphology and voice, Xenobatrachus lanthanites sp. nov. has the greatest similarity to Xenorhina bouwensi , but the latter lacks palatovomerine spikes.
Description of the holotype. Head extremely small, broader than long and merging seamlessly into the wider body; snout acuminate from above ( Fig. 5 View FIGURE 5 a), protruding in profile ( Fig. 5 View FIGURE 5 b), loreal region oblique, no canthus rostralis, nostrils visible from above and dorsolaterally directed, internarial distance about the same as eye diameter and distance between eye and naris. Tympanum scarcely visible and very large (94% of eye diameter), weak supratympanic fold (more conspicuous in life). Legs short, no webs between fingers or toes, fingers rather wide, their tips not wider than terminal phalanges and with hardly visible grooves, toe tips a bit wider than terminal phalanges and with clearly marked terminal grooves, relative length of fingers 3>4>2>1 ( Fig. 6 View FIGURE 6 a), relative length of toes 4>3>5>2>1( Fig. 6 View FIGURE 6 b), no subarticular tubercles, a few inconspicuous palmar and plantar tubercles. Dorsal and ventral surfaces in life and in preservative smooth, lateral surface in life with tubercles, in preservative smooth, tip of snout pustulose.
Colour in preservative: Dorsal surface greybrown, semicircular whitish spots surrounding blackish spots in the inguinal region (lumbar ocelli), snout whitish, extremities lighter than dorsum and with irregular dark spots and stripes, a large blackish triangular spot bordered above by a whitish line around cloaca, a blackish stripe also from posterior eye angle to insertion of fore limb, ventral surfaces crème coloured and densely reticulated with brownish spots.
Colour in life: Dorsal surface dark brown with diffuse small blackish spots, snout tip whitish, dorsal surface of distal part of tibiae and of entire tarsi conspicuously brightened, light vertebral line and supratympanic fold more pronounced than in fixative. Iris nearly circular, dark brown and with golden inner margin. Ventral surface of the same colour as in fixative.
Morphological variation in the type series. Variation in biometric characters is fairly low ( Table 2 View TABLE 2 ). SUL varies only between 21.3 and 22.4 mm (mean 21.9 mm) and TL/SUL from 0.40–0.42 (mean 0.41) in five adult males. Colouration in preservative varies as follows (only four specimens are considered): two specimens have a light greybrown dorsal surface of head, body and extremities with a few irregular dark brown small spots and stripes; the other two are dark grey with a very indistinct mottling; lumbar ocelli are clearly expressed in two specimens, only indicated in the other two; there is a fine light vertebral line in two specimens and dark stripes and/or spots around the tympanum in all specimens; all specimens have a large triangular blackish spot, bordered by a whitish stripe above, around the anal opening; all specimens have a depigmented tip of the mouth, which in three specimens exhibits dark pustules; distal part of the tibiae and part of the tarsi much lighter than the remaining dorsal surface in one specimen (this was the case at least in two specimens when they were alive); belly and lower flanks are strongly mottled in one and nearly unspotted in another specimen but slightly so in the others; mottling on the chin is well expressed in all specimens. Basic colour in life varied from different tones of brown to different hues of bluegrey. Clearly visible pustules on the body sides were striking in the living specimens, but disappeared nearly completely in fixative.
Osteological characteristics (based on the cleared and double stained paratype ZMB 69561). Most skull bones robust. Frontoparietals and nasals with a median suture and in close contact to one another; a large squamosal, its otic ramus strongly overlapping the crista parotica, anterior ends of maxillaries nearly meet (but do not fuse) in front of the premaxillaries ( Fig. 7 View FIGURE 7 a); dentary and angular bone strongly developed, anterior ends of dentaries left a wide chasm in front of the mentomeckelian bones, quadratojugal firmly attached to dentary. Prevomer and palatine fused to a vomeropalatine and in firm contact mesially with its neighbour, these vomeropalatines are strongly extended anteriorly and posteriorly and each bear a large posteriorly and inferiorly directed spike ( Fig. 7 View FIGURE 7 b). Hyoid plate with cartilaginous anterolateral and posterolateral processes and very long ossified posteromedial processes; the latter bear posteriorly directed processes in the middle of their anterior sides ( Fig. 7 View FIGURE 7 c). Clavicles, procoracoids and omosternum absent. Coracoids strongly developed and fused to scapulae, their median ends much wider than the distal ones; sternum cartilaginous and with a wide xiphisternum ( Fig. 7 View FIGURE 7 d). Eight presacral vertebrae; the first with “unusually” long anterolateral processes; relative width of transverse processes: sacrum>IV>II>V>VI=VII=VIII; vertebrae I–IV with neural spines of different size, sacral diapophyses barely dilated. A bicondylar sacralurostyle articulation, ilia and urostyle with dorsal crests ( Fig. 7 View FIGURE 7 e). Phalangeal formula of hand 2, 2, 3, 3, no intercalary structures, terminal phalanges end with no or only very small transverse processes. Prepollex cosists of two very small cartilaginous elements. The preparation has only one malformed foot, therefore no statements can be made concerning foot bones.
Habitat and habits. All frogs were found near a road traversing a partly logged mossy rain forest at an altitude of about 1000 m a.s.l. ( Fig. 8 View FIGURE 8 ). The frogs started calling during twilight and called with shorter or longer breaks until midnight. Their calling sites were in small holes 5–20 cm below the surface, beneath trees or shrubs. Most animals occurred in loose groups. Minimal distance between two calling males was two meters. Interestingly, no antiphonal duets were heard (in contrast to Xenorhina bouwensi ). Xb. lanthanites sp. nov. lives syntopically with Xenorhina varia , two Platymantis , one Choerophryne , four Oreophryne , one Hylophorbus , one Austrochaperina , one Cophixalus and three Litoria .
Vocalisation. The advertisement calls consist of series of low pitched piping notes, very similar to the calls of Xenorhina bouwensi . The apparently differences in the oscillograms and audiospectrograms, shown here ( Figs 2 View FIGURE 2 and 9 View FIGURE 9 ), are obviously caused by a different recording situation. The note series lasted from a few seconds to more than one minute. Note rate was from 1.2 to 1.8 notes/s. We found note lengths from 84 to 172 ms (mean 121 ms, SD 26.2) and internote lengths from 397–896 ms. Note length as well as internote length varied significantly between various specimens. For example, average interval length in one specimen was 447 ms, in another it was 621 ms and in a third frog it was 696 ms. Interval length of the first frog was identical to that of a Xenorhina bouwensi specimen from the Wondiwoi Mountains and note length of another specimen was also identical to that of a Xr. bouwensi . In general it can be stated that note and internote length of Xr. bouwensi are shorter than that of Xb. lanthanites sp. nov. and note rate consequently is higher in the former. Dominant frequency was around 1.0 kHz ( Fig. 10 View FIGURE 10 ), the same as in Xr. bouwensi .
Distribution. Known only from the type locality.
Etymology. The specific epithet lanthanites refers to the hidden lifestyle of the new species. Lanthan is the root of the Greek verb lanthanein meaning hidden or to hide and –ites is the Greek suffix indicating “having the nature of”. The specific epithet is masculine and matches the generic name Xenobatrachus in gender.
Molecular results. The amplified fragments of the partial mt 12S rRNA gene had a length of 650 bp in some samples but “only” 350 bp in others. Sequences of 350 bp of the following specimens were compared: Four specimens of Xenorhina bouwensi (ZMB 62638, 6513840), one specimen each of Xr. oxycephala (ZMB 69652) and Xr. varia (ZMB 65136), and two specimens of Xenobatrachus lanthanites sp. nov. (ZMB 69559, 69561). Uncorrected genetic distances (‘pdistances’) between the different sequences are listed in table 3.
Intraspecific genetic distances are much lower in comparison with interspecific distances. The genetic distances between the different species lie within a range of 4.5 % to 13 % with Xr. varia and Xr. oxycephala showing the closest genetic similarity.
Comparison with other species. Overall habitus and presence of toothlike spikes on the vomeropalatines ( Fig. 7 View FIGURE 7 b) clearly allocates the new species to the genus Xenobatrachus . Adult specimens of most of the 18 known species of this genus have a SVL of more than 30 mm ( Zweifel 1972, Menzies & Tyler 1977, Blum & Menzies 1989, Kraus & Allison 2002). Of the smaller species, Xb. ophiodon and Xenobatrachus bidens (van Kampen) have two instead of one spike on each vomeropalatine. Differences to Xenobatrachus anorbis were pointed out in the paragraph “Diagnosis”. Xb. lanthanites sp. nov. differs from Xenobatrachus schiefenhoeveli Blum & Menzies by longer tibiae (TL/ SUL(Zw) 0.40–0.42 vs. 0.31–0.36) and larger eyes (ED/SUL 0.071–0.081 vs. 0.065–0.074), moreover, the latter occurs in open grassland and never in forests (Blum & Menzies 1988); from Xenobatrachus tumulus Blum & Menzies by longer tibiae (TL/ SUL(Zw) 0.40–0.42 vs 0.38–0.39), larger eyes (ED/SUL 0.071–0.081 vs. 0.058–0.074) and a different ratio END / IND ( 0.94–1.20 in Xb. lanthanites sp. nov. and 1.11–1.28 in Xb. tumulus ); moreover, advertisement calls of Xb. tumulus consist of less than 20 notes with note lengths of 60–70 ms, whereas calls of Xb. lanthanites sp. nov. have much more than 20 notes and note duration is 100 ms or more. Xb. lanthanites sp. nov. may be distinguished from Xenobatrachus mehely (Boulenger) mainly by its smaller size and the ratio END / IND (mean in Xb. mehely 1.31 according to Zweifel, 1972, and in Xb. lanthanites sp. nov. 1.11). Moreover, Xb. mehely calls contain less than 20 notes, internote intervals last about 1500 ms and note repetition rate is less than one per second (Blum & Menzies 1988). Recall that Xb. lanthanites sp. nov. utters much more notes per call with internote intervals of less than 800 ms and a note repetition rate of 1–2/s. Xenobatrachus subcroceus Menzies & Tyler measures from 30.5–33.3 mm SVL (n=7) and thus has a much greater body size than Xb. lanthanites sp. nov. Most data on the compared species were obtained from the papers of the abovementioned authors.
As already pointed out, the new species has great similarities in external morphology, anatomy, ecology and advertisement calls to Xenorhina bouwensi . The two species could easily be confused at first glance. The best diagnostic trait to distinguish them is the presence of vomeropalatine spikes in Xb. lanthanites sp. nov. which are lacking in Xr. bouwensi . Other differences are in body size, in the ratios END / IND and T4D/F3D and in some call characteristics. Xb. lanthanites sp. nov. is significantly larger than Xb. bouwensi : SUL of the former ranges from 21.3–22.4 mm (n=5) and that of the latter (n=13) from 16.0–22.0 mm (tvalue of the means after Student 3.67, p=0.002). The ratio END / IND is in Xr. bouwensi 1.1–1.7 (mean 1.34) and that of Xb. lanthanites sp. nov. is from 0.9–1.2 (mean 1.11) (tvalue 2.74, p=0.0073). The ratio T4D/F3D is in Xr. bouwensi 1.7–2.7 (mean 2.01) and that of Xb. lanthanites sp. nov. is from 1.5–2.0 (mean 1.67) (tvalue 2.74, p=0.0075). Xr. bouwensi has a note repetition rate of 1.5–3.2 (mean 1.95) per second and Xb. lanthanites sp. nov. has a rate of 1.2–1.8 (mean 1.41) (tvalue 2.80, p=0.0078). The length of 115 notes from Xr. bouwensi varied from 79–104 ms (mean 89 ms) and that of 52 notes from Xb. lanthanites sp. nov. ranged between 84 and 172 ms (mean 121 ms) (tvalue 12.36, p=2.4 X 107). 40 internote intervals from Xr. bouwensi were from 397–598 ms (mean 447 ms) and 35 internote intervals from Xb. lanthanites sp. nov. were from 480–797 ms (mean 621 ms) (tvalue 11.27).
There are some more minor differences between Xb. lanthanites sp. nov. and Xr. bouwensi : fingers and toes of the former are wider and have a better developed callus internus, Xr. bouwensi has two short longitudinal skin ridges in the shoulder region (lacking in Xb. lanthanites sp. nov.), and its skeleton is more lightlybuilt.
Concluding remarks. Xenobatrachus lanthanites sp. nov. exhibits greater similarities in external morphology, anatomy, ecology and behaviour to Xenorhina bouwensi than to any species within its own genus.
The only remarkable anatomical trait to distinguish between both species and the genera Xenobatrachus and Xenorhina is the presence of toothlike spikes on the vomeropalatine in the former. This fact was also stressed by Zweifel (1972), Burton (1986) and Menzies & Blum (1989). The question arose whether both genera should be maintained or whether Xenobatrachus should be treated as a junior synonym of Xenorhina .
We tried to help solve this question by means of molecular investigations.
Comparisons of sequences of 350 bp of the mt 12S rRNA gene from Xb. lanthanites sp. nov. and three species of Xenorhina showed a great genetic distance (13 %) between Xb. lanthanites sp. nov. and Xr. bouwensi (it should be stressed here that our studies of the same sequences in the welldifferentiated European brown frogs Rana arvalis Nilsson and Rana dalmatina Fitzinger led to a genetic distance of “only” 4 %). However, genetic distances between Xr. bouwensi and Xr. oxycephala as well as Xr. varia are of a similar level ( Table 3 View TABLE 3 ). Considering these biochemical results, it would be premature to synonymize Xenobatrachus with Xenorhina . In contrast, the overall similarities of Xb. lanthanites sp. nov. and Xr. bouwensi in the light of genetic studies seem more likely a result of parallel evolution than of “true” kinship. Both species match the criteria of “sibling species” as expressed by Mayr (1975), but they are probably not sister taxa in terms of phylogentic systematics. More detailed biochemical studies (investigations of longer sequences of various genes of more species in both genera) and investigations of other characteristics are necessary to solve the problem whether Xenobatrachus should be regarded as a junior synonym of Xenorhina .
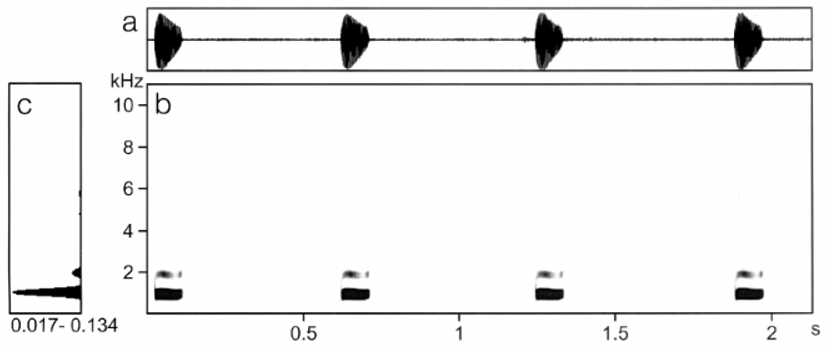



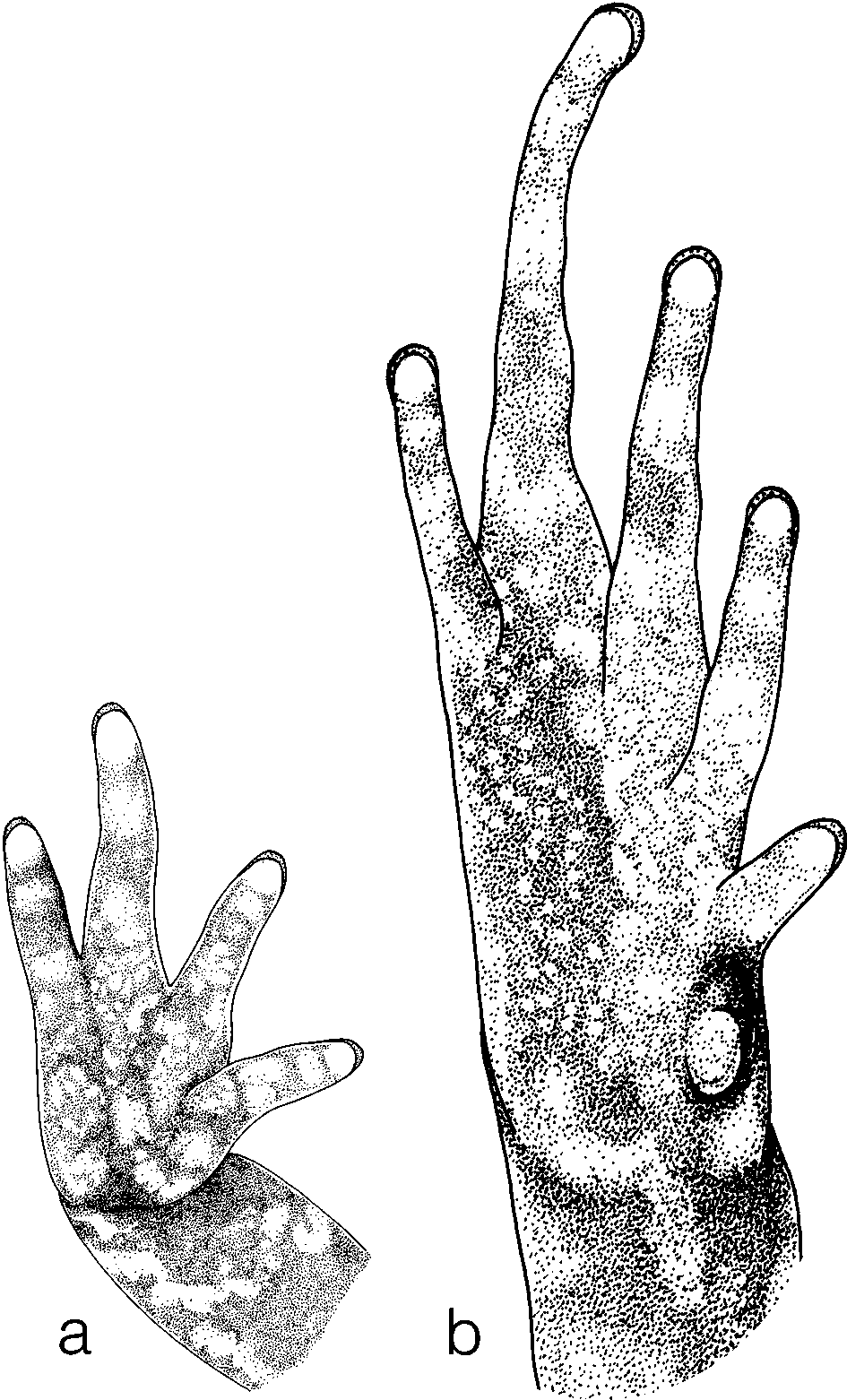
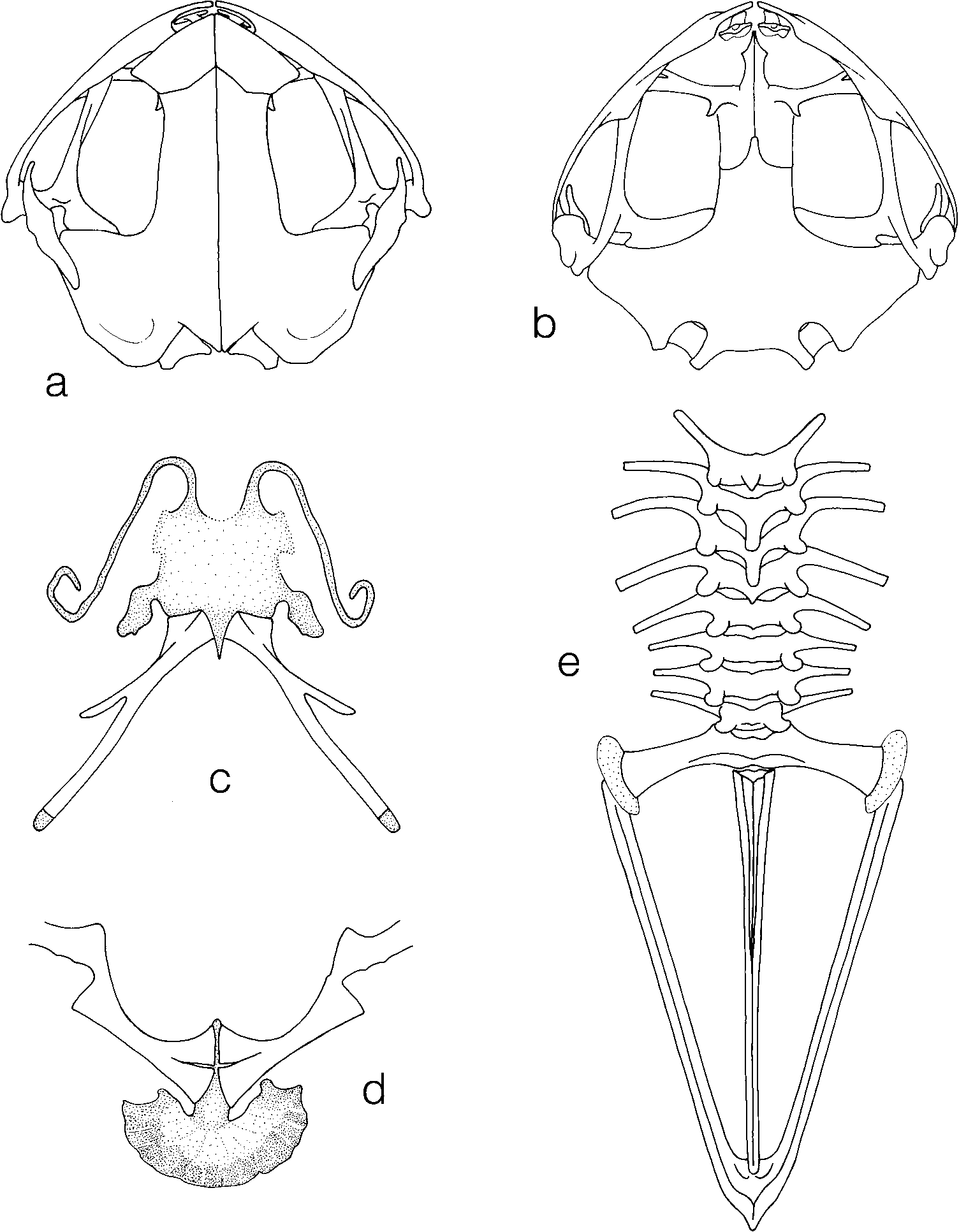

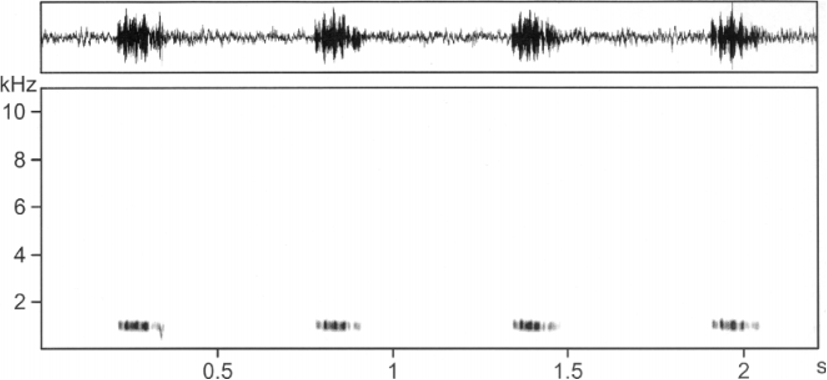
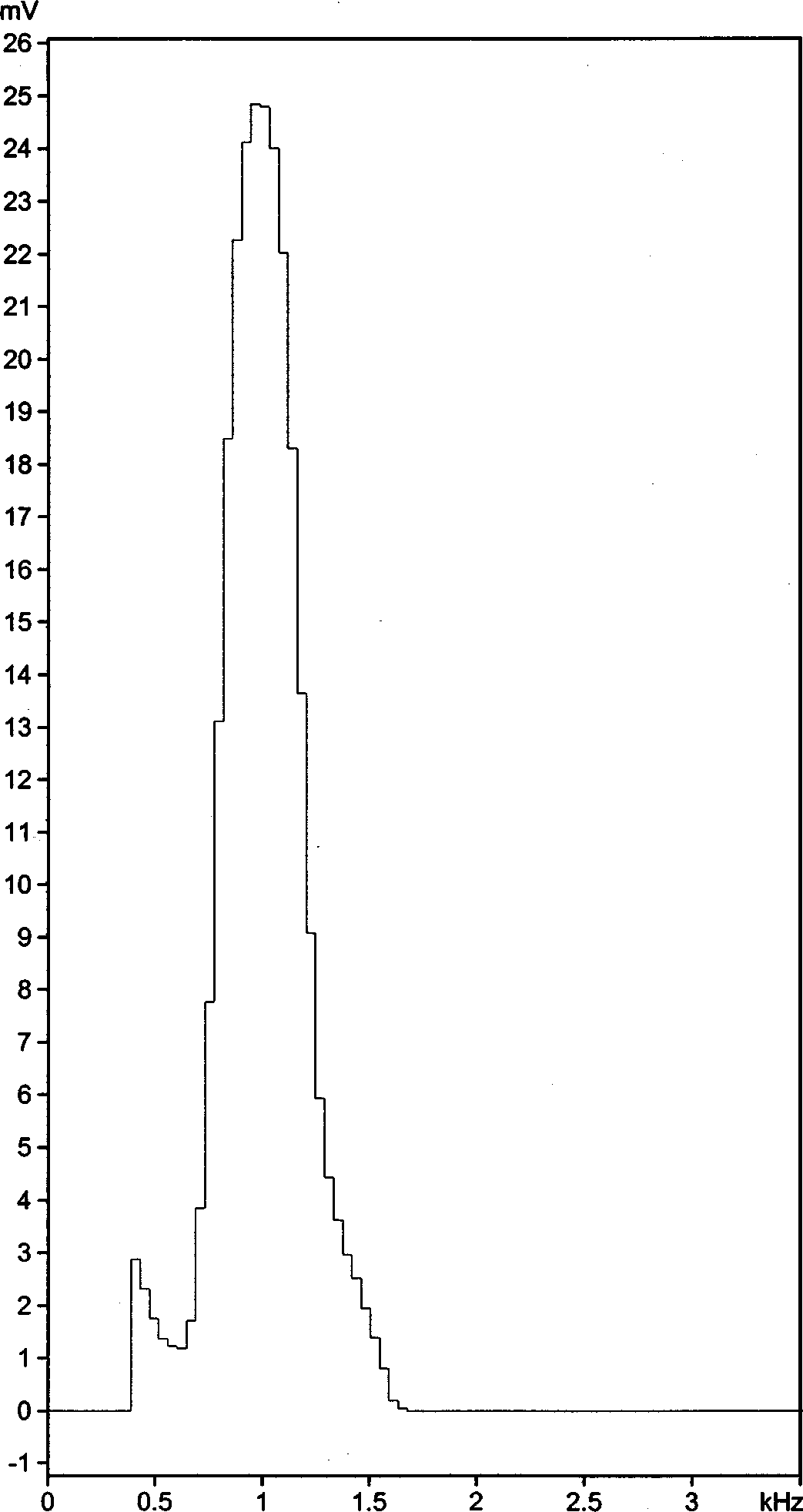
TABLE 2. Biometric values of the type series of Xenobatrachus lanthanites sp. nov., all measurements in mm. Abbreviations are explained in the paragraph ” Material and methods ”; ZMB 69558 is the holotype and ZMB 69561 was cleared and stained as an osteological preparation.
| ZMBNo. 69557 | 69558 | 69559 | 69560 | 69561 |
|---|---|---|---|---|
| FN 7787 | 7788 | 7789 | 7790 | 7791 |
| SUL 21.3 | 22.4 | 21.4 | 22.2 | 22.0 |
| TL(Zw) 9.0 | 8.9 | 8.7 | 9.1 | 8.9 |
| TL(RG) 9.8 | 10.1 | 9.6 | 9.8 | 10.0 |
| TaL 7.1 | 6.5 | 6.4 | 6.5 | 6.4 |
| L4T 9.4 | 9.0 | 9.3 | 9.5 | |
| L1T 1.9 | 1.5 | 1.7 | 1.6 | |
| LMT 0.5 | 0.5 | 0.7 | 0.7 | |
| T4D 0.7 | 0.8 | 0.6 | 0.6 | 0.8 |
| F3D 0.4 | 0.5 | 0.4 | 0.4 | 0.4 |
| HL 5.8 | 6.5 | 6.4 | 6.5 | 6.3 |
| HW 7.9 | 7.8 | 7.7 | 7.8 | 8.0 |
| END 1.6 | 1.8 | 1.8 | 1.7 | 1.8 |
| IND 1.7 | 1.5 | 1.5 | 1.5 | 1.7 |
| ED 1.7 | 1.6 | 1.7 | 1.8 | 1.6 |
| TyD 1.5 | 1.5 | 1.5 | 1.4 | 1.4 |
| SL 2.8 | 2.7 | 2.8 | 2.8 | 2.5 |
| OD 5.8 | 5.0 | 4.6 | 4.9 | 4.9 |
| TL/SUL(Zw) 0.42 | 0.40 | 0.41 | 0.41 | 0.40 |
| TL/SUL(RG) 0.46 | 0.45 | 0.45 | 0.44 | 0.45 |
| TaL/SUL 0.33 | 0.29 | 0.30 | 0.29 | 0.29 |
| T4D/SUL 0.033 | 0.036 | 0.028 | 0.027 | 0.036 |
| T4D/F3D 1.75 | 1.60 | 1.50 | 1.50 | 2.00 |
| MTL/T1L 0.26 | 0.30 | 0.41 | 0.44 | |
| HL/SUL 0.27 | 0.29 | 0.30 | 0.29 | 0.29 |
| HL/HW 0.73 | 0.83 | 0.83 | 0.83 | 0.79 |
| END/IND 0.94 | 1.20 | 1.20 | 1.13 | 1.06 |
| ED/SUL 0.080 | 0.071 | 0.079 | 0.081 | 0.073 |
| TyD/ED 0.88 | 0.94 | 0.88 | 0.78 | 0.88 |
| SL/SUL 0.13 | 0.12 | 0.13 | 0.13 | 0.11 |
| OD/SUL 0.27 | 0.22 | 0.21 | 0.22 | 0.22 |
TABLE 3. Uncorrected genetic distances (“ pdistances ”) between the sequences of Xenobatrachus lanthanites sp. nov. and various Xenorhina species.
| Xenorhina | Xenobatrachus | |||||
|---|---|---|---|---|---|---|
| bouwensi | oxycephala | varia | lanthanites sp. nov. | |||
| Xenorhina | ZMB ZMB 65138 65139 | ZMB 65140 | ZMB 69652 | ZMB 65136 | ZMB ZMB 69559 69561 | |
| bouwensi | ZMB 62638 | <3 % <3 % | <3 % | 13 % | 12 % | 13 % 13 % |
| ZMB 65138 | <3% | <3 % | 13 % | 12 % | 13 % 13 % | |
| ZMB 65139 | <3 % | 13 % | 12 % | 13 % 13 % |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |