
Echinoderes juliae, Sørensen & Rohal & Thistle, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.456 |
|
publication LSID |
lsid:zoobank.org:pub:DE1B1DEE-9871-4803-9F67-025F2B439560 |
|
DOI |
https://doi.org/10.5281/zenodo.3818850 |
|
persistent identifier |
https://treatment.plazi.org/id/3235E6EC-DA74-4269-8251-28654AA0EE69 |
|
taxon LSID |
lsid:zoobank.org:act:3235E6EC-DA74-4269-8251-28654AA0EE69 |
|
treatment provided by |
Valdenar |
|
scientific name |
Echinoderes juliae |
| status |
sp. nov. |
Echinoderes juliae sp. nov.
urn:lsid:zoobank.org:act:3235E6EC-DA74-4269-8251-28654AA0EE69
Figs 14–17 View Fig View Fig , Tables 10–11
Diagnosis
Echinoderes with middorsal spines on segments 4 to 8, and spines in lateroventral positions on segments 6 to 9. Tubes present in lateroventral positions on segment 5. Glandular cell outlets type 2 present in subdorsal, laterodorsal, sublateral and ventrolateral positions on segment 2, in sublateral positions on segments 3, 4 and 8, and in midlateral positions on segment 5. Segment 11 with small, middorsal protuberance. At least posterior half of tergal plate of segment 11 divided by longitudinal fissure. Males with three pairs of penile spines; females with long lateral terminal accessory spines.
Etymology
The first author (MVS) dedicates this species to his friend and former meiofauna colleague Julia Zekely.
Material examined
Holotype
UNITED STATES OF AMERICA: adult ♀, US West Coast, California, off Monterey , 36°40′52″ N, 122°49′37″ W, St. 6, 2719 m deep, collected from mud, mounted in Fluoromount G on a glass slide, 24 Sep. 2008 (NHMD-213666). See Fig. 1 View Fig for localities and Table 1 View Table 1 for detailed station data.
GoogleMapsParatypes
UNITED STATES OF AMERICA: 4 ♀♀, 5 ♂♂, same collecting data as for holotype; 1 ♂, St. 2; 1 ♀, St. 3; 1 ♀, 1 ♂, St. 4; 2 ♀♀, 1 ♂, St. 5; 1 ♀, 1 ♂, St. 8 (NHMD-213667–213691). All paratypes are mounted in Fluoromount G, one on H-S slide and the remaining on glass slides.
Additional non-type material
UNITED STATES OF AMERICA: 4 ♀♀, 2 ♂♂, same collecting data as for holotype, mounted for SEM and stored in the first author’s personal reference collection.
Description
Adults with head, neck and eleven trunk segments ( Figs 14 View Fig A–B, 15, 17A–B). The trunk appears slender, with nearly parallel-sided segments in the anterior end, and slightly tapering trunk from segment 8 ( Fig. 15 View Fig A–B). Lateral terminal spines long and slender, around 75% of trunk length, and lateral terminal accessory spines in females long as well, around 60% of lateral terminal spine length ( Fig. 14 View Fig A–C). For complete overview of measurements and dimensions, see Table 10. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, is summarized in Table 11 View Table 11 .
All specimens had their heads completely retracted; hence, no details on head morphology can be provided. The neck has 16 placids, measuring 14 µm in length. The midventral placid is broadest, measuring 12 µm in width at its base, whereas all others are narrower, measuring 8 µm in width at their bases. Four dorsal and two ventral trichoscalid plates are present, with the two dorsal pairs being somewhat triangular, whereas the ventral pair is more rounded.
Segment 1 consists of a complete cuticular ring. Sensory spots are located near the anterior margin in subdorsal and laterodorsal positions ( Figs 14A View Fig , 16A, C); sensory spots are small and rounded, and lined with cuticular hairs. Glandular cell outlets type 1 present in middorsal and sublateral positions. Dorsal and lateral sides have scattered cuticular hairs, whereas the ventral side is devoid of hairs except for a single line on the posterior part of the segment. The posterior segment margin is nearly straight, terminating in a pectinate fringe with relatively short and pointed fringe tips.
Segment 2 consists of a complete cuticular ring. Pachycyclus of the anterior segment margin is relatively thin and not interrupted. Glandular cell outlets type 2 are present in subdorsal, laterodorsal, sublateral and ventrolateral positions ( Figs 14 View Fig A–B, 15, 16A–C, 17C–D). Sensory spots are located in middorsal ( Fig. 17C), laterodorsal, midlateral and ventromedial ( Fig. 17D) positions. Sensory spots are extremely minute, consisting of one or two pores and a few papillae (see Fig. 17C inset), but with rather long, thin marginal cuticular hairs; all sensory spots from this segment and to segment 10 have this appearance. Glandular cell outlet type 1 present in middorsal position. Secondary pectinate fringe present near anterior segment margin of this and the following segments, but it is usually covered by the preceding segment. On this and the eight following segments, the cuticular hairs are bracteate and fairly long, but not that dense, covering the dorsal and lateral parts of the segment. Posterior segment margin as on preceding segment.
Segment 3, and remaining segments, consist of one tergal and two sternal plates ( Figs 14B View Fig , 15B View Fig , 16B, E, H). Pachycyclus of the anterior segment margin is well-developed, and interrupted at the tergosternal and midsternal junctions and middorsally. Glandular cell outlets type 2 present in sublateral positions ( Figs 14B View Fig , 15C View Fig , 16C, 17F), sensory spots in subdorsal positions and glandular cell outlets type 1 in middorsal (Fig. 16C) and ventromedial (Fig. 16B) positions. Posterior segment margin nearly straight, with fringe tips being slightly longer that those on preceding segment. Cuticular hairs cover the tergal plate and the lateral halves of the sternal plates.
Segment 4 with middorsal acicular spine ( Figs 14A View Fig , 16A) and glandular cell outlets type 2 present in sublateral positions ( Figs 14B View Fig , 15B View Fig , 16C, 17F–G). Sensory spots absent. Glandular cell outlets type 1 present in paradorsal (Fig. 16C) and ventromedial positions. Cuticular hairs basically as on preceding segment, except for the middorsal and paradorsal areas that are left hairless. Pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 5 with middorsal acicular spine ( Figs 14A View Fig , 16D), minute tubes in lateroventral positions ( Figs 14B View Fig , 16E, 17E, G), and glandular cell outlets type 2 present in midlateral positions ( Figs 14 View Fig A–B, 15C, 16C, 17G). Sensory spots present in subdorsal (Fig. 16D) and ventromedial ( Fig. 17E) positions. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin, and cuticular hairs as on preceding segment.
Segment 6 with acicular spines in middorsal and lateroventral positions ( Figs 14 View Fig A–B, 16D–E, 17E, G). Sensory spots present in paradorsal, subdorsal (Fig. 16D), midlateral and ventromedial (Figs 16E, 17E) positions. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Fig. 16 (opposite page). Light micrographs showing overviews and details of Echinoderes juliae sp. nov., holotype, ♀ (A–B, D–E, G–H) (NHMD-213666), paratype, ♀ (C) (NHMD-213689), paratype, ♂ (F) (NHMD-213686) and paratype, ♀ (I) (NHMD-213684). A. Segments 1 to 5, dorsal view. B. Segments 1 to 5, ventral view. C. Segments 1 to 6, lateral view. D. Segments 5 to 7, dorsal view. E. Segments 5 to 7, ventral view. F. ♂, segments 10 to 11, ventral view. G. Segments 8 to 10, dorsal view. H. Segments 8 to 10, ventral view. I. ♀, segments 10 to 11, ventral view.Abbreviations: ldgco2 = laterodorsal glandular cell outlet type 2; ldss = laterodorsal sensory spot; ltas = lateral terminal accessory spine; lts = lateral terminal spine; lvs = lateroventral spine; lvt = lateroventral tube; mdgco1 = middorsal glandular cell outlet type 1; mds = middorsal spine; mlgco2 = midlateral glandular cell outlet type 2; pdgco1 = paradorsal glandular cell outlet type 1; pdss = paradorsal sensory spot; pe = penile spines; pvgco1 = paraventral glandular cell outlet type 1; sdgco2 = subdorsal glandular cell outlet type 2; sdss = subdorsal sensory spot; slgco2 = sublateral glandular cell outlet type 2; te = tergal extensions; vlgco2 = ventrolateral glandular cell outlet type 2; vlss = ventrolateral sensory spot; vmgco1 = ventromedial glandular cell outlet type 1; vmss = ventromedial sensory spot.
Segment 7 with acicular spines in middorsal and lateroventral positions ( Figs 14 View Fig A–B, 16D, 17H). Sensory spots present in paradorsal, midlateral and ventromedial positions; ventromedial sensory spots slightly more lateral than on the two preceding segments. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 8 with acicular spines in middorsal and lateroventral positions ( Figs 14 View Fig A–B, 16G–H, 17H), and large glandular cell outlets type 2 present in sublateral positions ( Figs 14B View Fig , 15 View Fig B–C, 16H, 17H). Sensory spots present in paradorsal positions only (Fig. 16G). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 9 with acicular spines in lateroventral positions ( Figs 14B View Fig , 16H). Sensory pots present in paradorsal, subdorsal (Figs 16G, 17I), midlateral and ventrolateral (Fig. 16H) positions. Small rounded sieve plates are present in sublateral positions ( Fig. 14B View Fig ). Cuticular hair covering on tergal plate even less dense than on preceding segments, and subdorsal to middorsal areas only with a few scattered hairs anteriorly on segment ( Fig. 17I). Glandular cell outlets type 1, pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 10 without spines or tubes. The pectinate fringe of the posterior segment margin is interrupted and has small indentations in the laterodorsal positions where tubes often occur in species of Echinoderes , but tubes are not present in this species ( Fig. 17K). Sensory spots are present in subdorsal and ventrolateral positions. Glandular cell outlets type 1 are present as two middorsal ones (Fig. 16G), and a pair in paraventral positions (Fig. 16H). A few cuticular hairs are scattered over the tergal plate and lateral parts of the sternal plates. The posterior segment margin of the tergal plate is straight, with shorter fringe tips. The margins of the sternal plates are broadly concave, also with slightly shorter fringe tips. Pachycycli as on preceding segment.
Segment 11 with lateral terminal spines ( Figs 14C View Fig , 17B). Males with three pairs of penile spines ( Figs 14 View Fig D–E, 16F, 17K); dorsal and ventral penile spines are thin, flexible tubes, whereas the median one is thick, conical, and stout. Females thin and delicate, but also with rather long lateral terminal accessory spines ( Figs 14 View Fig A–C, 16I, 17J). A short middorsal protuberance extends from the intersegmentary joint; the protuberance is flanked by numerous minute hairs and a pair of paradorsal sensory spots ( Figs 14A, D View Fig , 17 J–K). Glandular cell outlets type 1 present as two middorsal ones, in association with, or underneath, the protuberance. The segment is completely devoid of cuticular hairs, besides the short hair-like extensions around the protuberance ( Fig. 17J). The posterior part of the tergal plate appears to be divided by a longitudinal fissure ( Fig. 17J). Because of the overlying protuberance, it is difficult to visualize how far anterior the fissure expands, but the tergal plate can at least be considered as partially divided. Tergal extensions are long and slender, with a small tooth on the inferior margin, and extending into fairly long, flexible tips ( Figs 14 View Fig , 16F, I). Sternal extensions shorter, but also with a short projecting tooth.
Remarks
Despite its quite typical spine formula, the distribution of glandular cell outlets type 2, combined with its slender appearance, long lateral terminal accessory spines and conspicuously shaped tergal extensions, make it very easy to distinguish E. juliae sp. nov. from all other known species. Echinoderes juliae sp. nov. can be distinguished from other congeners by the presence of glandular cell outlets type 2 in sublateral positions on segments 3 and 4. No other known species have sublateral outlets on these segments.
Eight additional species have the combination of glandular cell outlets in subdorsal, laterodorsal, sublateral and lateroventral/ventrolateral positions on segment 2. These include E. angustus Higgins & Kristensen, 1988 , E. anniae sp. nov., E. cernunnos Sørensen et al., 2012 , E. drogoni , E. obtuspinosus Sørensen et al., 2012 , E. romanoi Landers & Sørensen, 2016 , E. tubilak Higgins & Kristensen, 1988 , and E. unispinosus (see Higgins & Kristensen 1988; Sørensen et al. 2012; Landers & Sørensen 2016; Grzelak & Sørensen 2018; Yamasaki et al. 2018b). All of these species also have glandular cell outlets type 2 in sublateral positions on segment 8, and five of them have midlateral glandular cell outlets type 2 on segment 5 as well. The latter are missing in E. obtuspinosus , E. anniae sp. nov. and E. unispinosus though, and none of these species can really be confused with E. juliae sp. nov. Also, the remaining five species differ in general appearance. E. angustus and E. tubilak furthermore differ from E. juliae sp. nov. by their having sublateral glandular cell outlets type 2 on segment 4 ( Grzelak & Sørensen 2018). Echinoderes drogoni has its tubes on segment 5 placed in lateral accessory positions ( Grzelak & Sørensen 2018), E. cernunnos has midlateral glandular cell outlets type 2 on segment 7 and very long, spinous tergal extensions ( Sørensen et al. 2012), and E. romanoi has much shorter tergal extensions and lateral terminal spines ( Landers & Sørensen 2016).
Echinoderes juliae sp. nov. probably shows the greatest overall resemblance with E. cernunnos and E. drogoni . Besides other similarities in spine and glandular cell outlet distribution, all three species share a rather rare trait, namely a full or partial longitudinal division of the tergal plate of segment 11. In E. drogoni the full division of the segment is evident in females only, but could be present in males as well. The males of E. drogoni , however, have a middorsal protuberance which, as is the case in E. juliae sp. nov., obscures the examination of the anterior part of the segment ( Grzelak & Sørensen 2018). The division is very clear in E. cernunnos , but only female morphology is known for this species ( Sørensen et al. 2012). In E. juliae sp. nov. the division is at least evident on the posterior half of the segment in both sexes. It is uncertain whether this suggests a closer relationship between the three species, but it is at least noteworthy that these, otherwise fairly similar species, also are the only three species of Echinoderes with full or partial tergal division of segment 11.
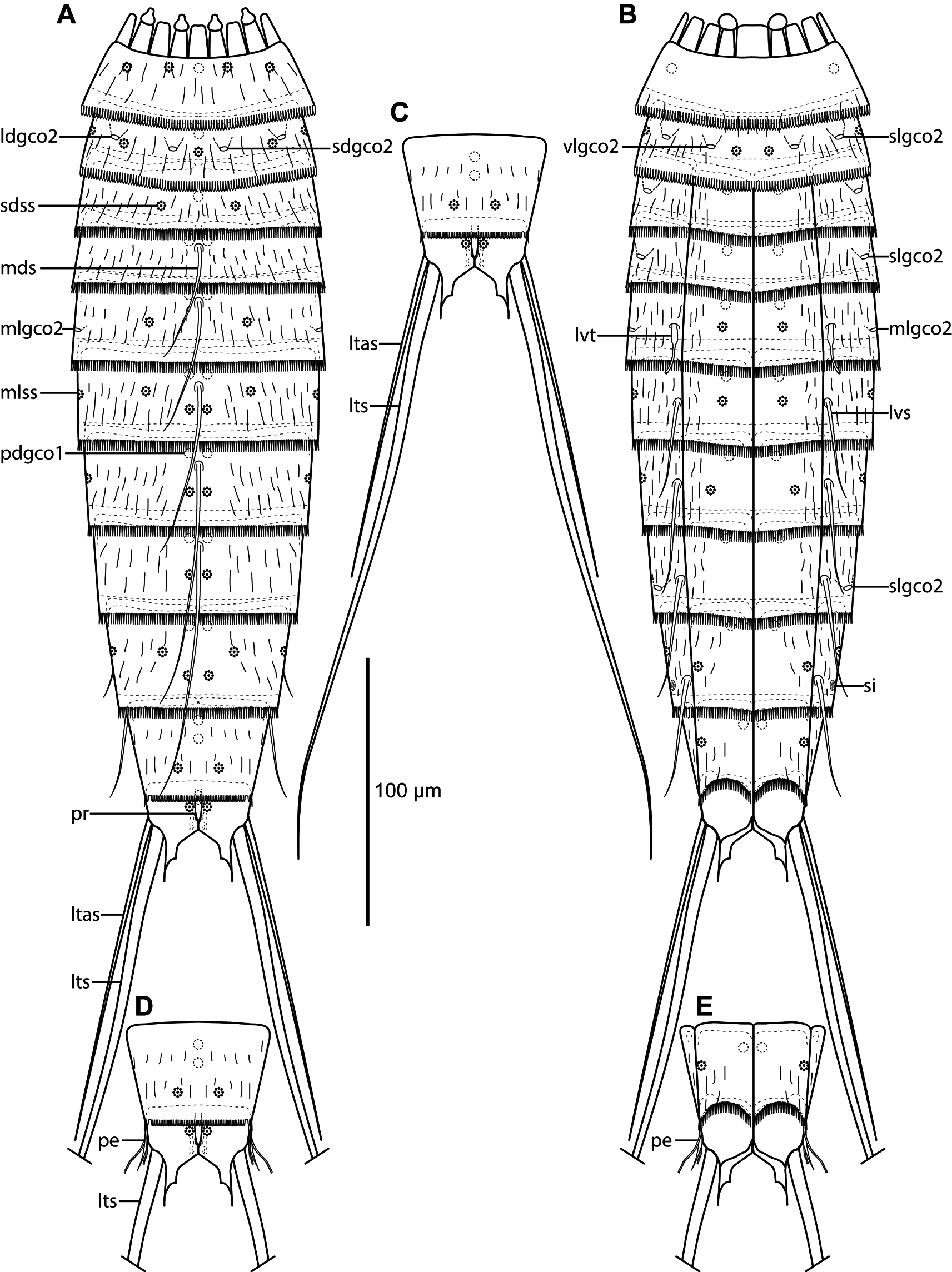

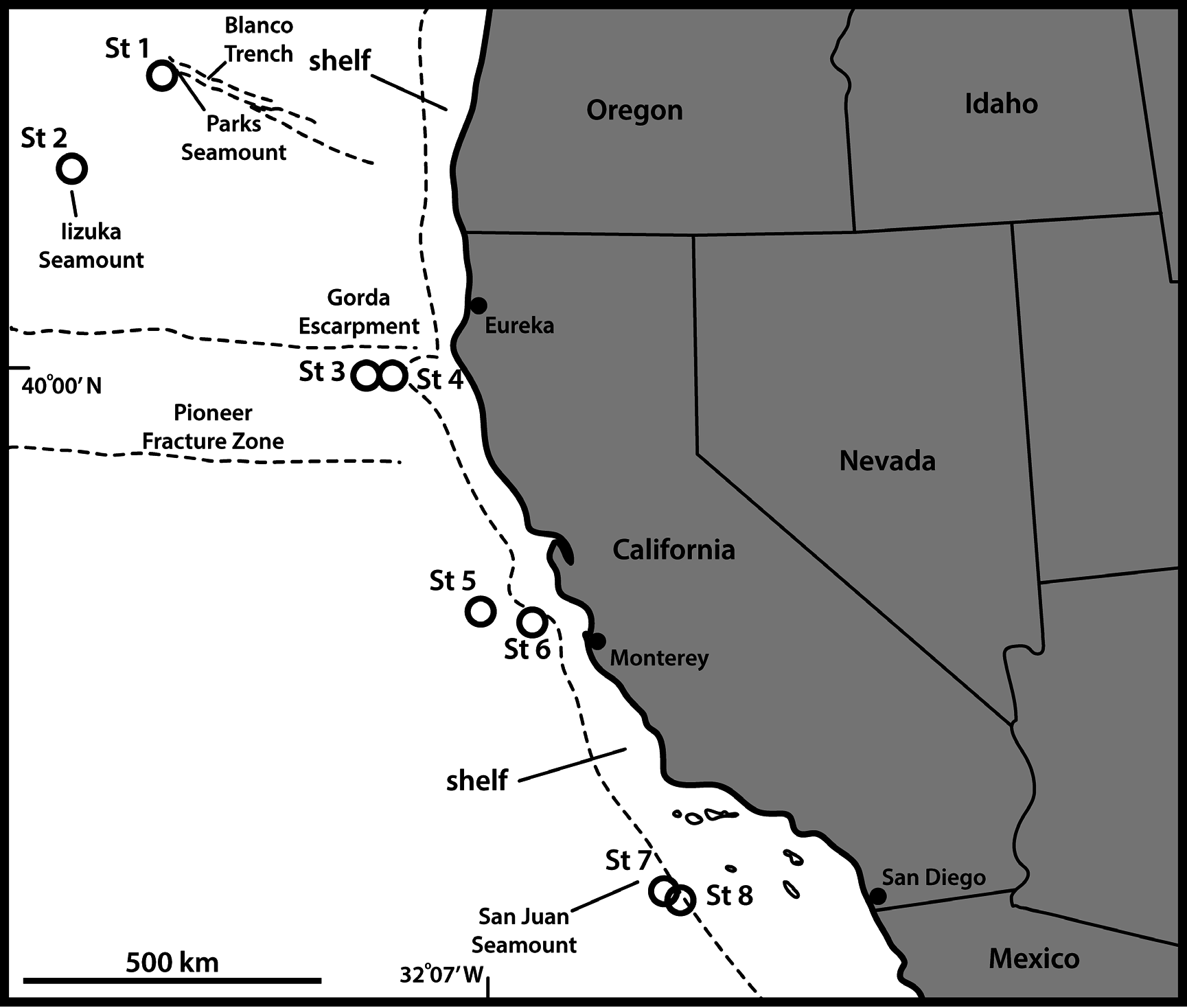
Table 11. Summary of nature and location of sensory spots, glandular cell outlets, tubes and spines arranged by series in Echinoderes juliae sp. nov. Abbreviations: LA = lateral accessory; LD = laterodorsal; LV = lateroventral; MD = middorsal; ML = midlateral; PD = paradorsal; PV = paraventral; SD = subdorsal; SL = sublateral; VL = ventrolateral; VM = ventromedial; ac = acicular spine; gco1/2 = glandular cell outlet type 1/2; ltas = lateral terminal accessory spine; lts = lateral terminal spine; pe = penile spines; pr = protuberance; si = sieve plate; ss = sensory spot; tu = tube; (♀) = female and (♂) = male conditions of sexually dimorphic characters.
| Position | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Segment MD | PD | SD | LD | ML | SL | LA | LV | VL | VM | PV | |
| 1 | gco1 | ss | ss | gco1 | |||||||
| 2 | gco1, ss | gco2 | ss, gco2 ss | gco2 | gco2 | ss | |||||
| 3 | gco1 | ss | gco2 | gco1 | |||||||
| 4 | ac | gco1 | gco2 | gco1 | |||||||
| 5 | ac | gco1 | ss | gco2 | tu | ss, gco1 | |||||
| 6 | ac | ss, gco1 | ss | ss | ac | ss, gco1 | |||||
| 7 | ac | ss, gco1 | ss | ac | ss, gco1 | ||||||
| 8 | ac | ss, gco1 | gco2 | ac | gco1 | ||||||
| 9 | ss, gco1 | ss | ss | si | ac | ss | gco1 | ||||
| 10 | gco1, gco1 | ss | ss | gco1 | |||||||
| 11 | gco1, gco1, pr ss | 3 x pe(♂) | ltas(♀) | lts | |||||||
| US |
University of Stellenbosch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |