
MEGATHERIIDAE Gray, 1821
|
publication ID |
https://doi.org/ 10.5281/zenodo.4650583 |
|
persistent identifier |
https://treatment.plazi.org/id/F749921E-9613-FFCF-D699-B8E6FBA0FAD7 |
|
treatment provided by |
Felipe |
|
scientific name |
MEGATHERIIDAE Gray, 1821 |
| status |
|
Family MEGATHERIIDAE Gray, 1821 Subfamily MEGATHERIINAE Gray, 1821
Genus Megatherium Cuvier, 1796
Megatherium altiplanicum n. sp.
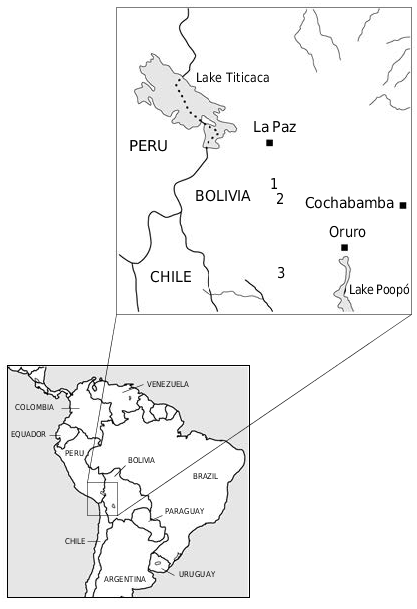
HYPODIGM. — Holotype ( MNHN AYO 101 ); this specimen preserves the palate, including both alveolar series, the maxillae anteriorly to the anterior root of the zygomatic arch; the horizontal ramus, angular process and part of the condyle of the left dentary; fragments of the horizontal and ascending rami, with parts of the coronoid process and condyle, of the left dentary; the atlas; two consecutive thoracic vertebrae and the neural arch of the next thoracic vertebra; the neural arches of three lumbar vertebrae; four consecutive caudal vertebrae; the proximal part of the right ulna; the left femur. TYPE LOCALITY. — The type specimen was recovered from near Ayo Ayo, Departamento La Paz, 17°05’S, 68°00’W, a village on the Altiplano. The fossiliferous locality is approximately 1 km south of Ayo Ayo. GoogleMaps
TYPE STRATA. — The Ayo Ayo-Viscachani fossiliferous locality ( Fig. 1 View FIG ) lies in the upper part of the Umala Formation.
ETYMOLOGY. — altiplanicum : neoformed Latin adjective (altiplanicus, - a, - um) meaning “relative to the Altiplano, from the Altiplano”, commemorating that the holotype and referred specimens derive from the Bolivian Altiplano.
REFERRED SPECIMENS. — From Ayo Ayo: Departamento La Paz, 17°05’S, 68°00’W: distal end of a left femur ( MNHN AYO 105 ) GoogleMaps ; right cuneiform ( MNHN AYO 150 ) ; left radius ( MNHN AYO 203 ) ; premaxillae of a juvenile individual ( MNHN AYO 230 ) ; proximal half of a right ulna ( MNHN BOL V 3304 ) ; right navicular ( MNHN BOL V 3312 ) .
From Viscachani: Departamento La Paz, approximately 17°09’S, 68°50’W: distal part of a left humerus (MNHN BOL V 3338).
From Pomata : Departamento Oruro, 18°20’S, 67°55’W: atlas ( MNHN POM 57 ) GoogleMaps ; posterior part of a right dentary, including angular process and condyle (MNHN BOL V 3294).
Fossils in MNHN were collected by Robert Hoffstetter in 1976. The type specimen was recovered from an area sufficiently restricted in size to permit its attribution to a single individual. Fossils in MNHN BOL were collected during collaborative field seasons between this institution and IFEA.
DIAGNOSIS. — Much smaller than M. americanum , and smaller also than M. medinae and M. tarijense . Ventral bulge of dentary relatively as deep as in M. americanum . Premaxillae firmly fused to each other and to maxillae, but less robust than in M. americanum . Palate narrows anterior to M1. Angular process distinct and more pointed than in M. americanum ; posterior part of the symphysis extends to the plane passing through m1. Femur twisted, with greater trochanter lying posterior to the plane passing through the head and distal condyles, lateral margin forming a sigmoid curve, and patellar trochlea reduced to the lateral half of the distal end of the femur, but apparently not to the degree as occurs in M. americanum .
GEOLOGY
The Umala Formation is bounded by the Toba 76 and Ayo Ayo volcanic tuffs, which lie at its base and top, respectively ( Fig. 2 View FIG ). Radiometric analyses of various minerals (i.e. biotite, sanidine) date the Toba 76 tuff at approximately 5.4 Ma, and the Ayo Ayo tuff at approximately 2.8 Ma, which indicate that the Umala Formation is of early and middle Pliocene age, according to the chronology of Marshall et al. (1992). M. altiplanicum n. sp. belongs to the Montehermosan Land Mammal Age.
The Pomata fossiliferous deposits ( Fig. 1 View FIG ) were initially referred to the Mauri Formation by Paz et al. (1966), but they are probably unrelated stratigraphically and ecologically to the Mauri Formation from the Oligocene and Miocene of the western Cordillera and western Altiplano. The Pomata locality lies above the conglomeratic facies of the Rosa Pata Formation, which is of Huayquerian age (“Conglomerado Pomata” of Marshall & Sempéré 1992), and above a volcanic tuff which probably represents a horizontal extension of the Toba 76 from the base of the Umala Formation.
DESCRIPTION
Skull
The type skull material ( Fig. 3 View FIG ) preserves the maxillae and premaxillae incompletely, and the right M1-M3 and left M1. Measurements are
Minimum palatal width
Between L and R M1 28.9
Between L and R M2 24.0
Between L and R M3 28.2
Between L and R M4 30.0
Midline palatal length, 143.0
level of M1 to choanae
Maximum height horizontal 145.0
ramus of dentary
Toothrow length between M1-M3 94.7
Lower toothrow length, alveolar 143.6
Maximum length and width of molariforms
M1 31.6; 29.1
M2 30.9; 36.4
M3 31.6; 35.0
m1 27.3; c. 34
m2 32.7; 34.0
m3 32.4; 30.0
m4 32.7; 25.5
given in Table 1. Laterally the right maxilla includes the anterior root of the zygomatic arch to the level of the buccal wall of the M1 alveolus. A smaller portion of the left maxilla is preserved to the level of the anterior root of the arch.
Premaxilla. The width across the premaxillae posteroventrally is comparable to the palatal interalveolar width (i.e. less than the width across the molariforms), which reflects the narrowed maxillae anterior to M1. Two large palatine fissures lie between and anterior to the M1s, approximately at the level where the palate narrows. The premaxillae are fused firmly to the maxillae. Their anterior portions are missing as they are broken a short distance anterior to the palatine fissures in MNHN AYO 101. Based on the preserved proximal portions, the premaxillae do not narrow abruptly anteriorly, which is also the condition in M. americanum , but not in Eremotherium laurillardi (Lund, 1842) and probably most other megatheriines ( De Iuliis 1994).
The complete premaxillae of a juvenile individual (MNHN AYO 230; Fig. 3A, B View FIG ) are unfused along their contact in the sagittal plane. In ventral view, each premaxilla is triangular, somewhat elongated, and the anterior extremity is almost imperceptibly widened. The distal, subterminal sagittal margins of the premaxillae are separated by a very flattened elliptical space. Anteriorly narrow, they widen progressively posteriorly, apparently following a constant radial curve. They are not rectangular, in contrast to the condition in adult individuals of M. americanum . The triangular condition, however, occurs in juvenile of the latter species (e.g., MACN 2830).
In M. altiplanicum n. sp. the ventral margin of the premaxilla is nearly flat in lateral view. The anterior third of the dorsal margin is dorsoventrally narrow; the central third rises at an angle of nearly 20°; the posterior third inclines slightly posteriorly. The dorsal surface bears a very obtuse angle between the posterior third and anterior two thirds. The posterior margin of the articulat- ed premaxillae is trapezoidal, with base ventral. The posterior surface of each premaxilla bears three processes, which were probably received by corresponding depressions in the maxilla. A ventromedial process joins that from the other premaxilla to form a ventrosagittal protuberance. A second process lies ventrolaterally. A third lies at the posterior extremity of the dorsolateral margin of the premaxilla.
Maxilla. In M. altiplanicum n. sp. the anterior margin of the anterior zygomatic root is level with a transverse plane passing through the middle of M1, as occurs in E. laurillardi ( Cartelle & De Iuliis 1995) , although it may lie opposite the septum between M1 and M 2 in the latter; in Megatheridium annectens Cabrera, 1928 , it reaches the mesial surface of M1. The root lies more posteriorly in M. americanum , in which its position varies from the septum between M1 and M2 to the distal part of M2. Apparently, the position is similar in M. medinae .
In ventral view the antemolariform region of the maxillar palate more closely resembles that in E. laurillardi , M. medinae and M. gallardoi , although to a lesser degree in the latter due to the brevity of this region, than in M. americanum ( Ameghino & Kraglievich 1921; Cabrera 1928; Hoffstetter 1952). In the first two species and M. altiplanicum n. sp., the antemolariform part of the maxillar palate is nearly as wide as the interalveolar width. Its lateral margins, between M1 and the premaxillae, are formed by « deux arêtes latérales qui se placent respectivement dans le prolongement de l’axe de chaque série dentaire », a condition considered primitive by Hoffstetter (1952: 60). In M. americanum and M. parodii Hoffstetter, 1949 , as well as in Plesiomegatherium halmyronomum Cabrera, 1928 , the antemolariform width of the maxillar palate is nearly equal to that between the buccal margins of the toothrows.
In M. altiplanicum n. sp., the palate is narrow between the toothrows, and its width is less than the transverse width of M1-M3, which is more similar to the condition in M. americanum , M. medinae , and Pyramiodontherium bergi (Moreno & Mercerat, 1891) than in E. laurillardi , Plesiomegatherium halmyronomum Cabrera, 1928 and Megatheridium annectens . In these last three, the palate width is about equal to or wider than the width of the largest molariform.
The palatine fissures lie between the premaxillomaxillary suture, approximately at the level where the premaxillae begin to narrow. Each fissure forms a shallow depression; its floor is pierced by a posterolateral foramen and a smaller, more anterior foramen.
Upper dentition. The upper molariforms ( Fig. 3D, E View FIG ) resemble those of other megatheriines. They are prismatic, quadrangular in section with width slightly exceeding length, and bear two transverse crests separated by a transverse, V-shaped valley.
M1 is less triangular in cross section than that of M. medinae and M. gallardoi (judging from the alveoli in the latter; Ameghino & Kraglievich 1921). Indeed, it resembles the M1 of M. americanum (as illustrated, for example, by Owen 1856), which is nearly trapezoidal in section, with a wider distal surface and a slightly rounded mesial surface. However, the shape of M1 varies between nearly trapezoidal to triangular in both M. americanum and E. laurillardi . The distal transverse crest is wider than the mesial, and its distal wear facet is considerably more prominent than the mesial facet of the mesial crest.
The mesial crest of M2 is slightly shorter transversely than the distal crest. The mesial wear facet of the mesial crest is more prominent than the distal facet of the distal crest. This also occurs in M3. The transverse crests of this molariform are nearly equal in transverse width. The crests of M2 and M3 are nearly rectilinear, but slightly convex mesially.
Dentary. The dentary of M. altiplanicum n. sp. is incompletely known. The holotype ( Figs 4 View FIG ; 5 View FIG ) preserves a right horizontal ramus, including right and left symphyseal regions as far as the mesial wall of the left m1 alveolus; a medial fragment of the right condyle; the right angular process; a lateral fragment of the alveolar region of the left horizontal ramus, including part of the anterior margin of the ascending ramus; a fragment of the ventral margin of the left horizontal ramus; and part of the left coronoid process, including the condyle.
The symphysis ends posteriorly at the level of the distal margin of m1. As in all megatheriines, the symphysis forms a trough-like spout. In dorsal view the lateral margins of the spout are parallel and extend anteriorly from the lingual half of the mesial margin of the m1 alveolus; this morphology reflects a narrowing of the lateral margin of the horizontal ramus. Further, the anterior margin of the spout is not rectilinear, but bears a distinct median notch. In lateral view, the dorsal margin of the spout continues anteriorly from the alveolar margin, but is not rectilinear. Its proximal two fifths are concave, its distal three fifths slightly convex. A mental foramen, representing the anterior emergence of the mandibular canal, lies on the lateral surface of the central part of the spout. The canal continues posteriorly within the lingual wall of the dentary. It emerges posteriorly, facing anteriorly and dorsally, between m4 and the anterior margin of the ascending ramus.
The ventral margin of the dentary of M. altiplanicum n. sp. bulges prominently, with maximum depth occurring beneath m3. The depth of the dentary relative to the toothrow length is nearly equal to that of M. americanum , in which depth exceeds that of all other megatheriines ( Kraglievich 1930; Hoffstetter 1952; Cartelle 1992) due to increased hypsodonty. The ventral margin of the dentary of M. lundi seijoi ( Kraglievich 1931: figs 1, 2) is more gently curved between the anterior extremity of the spout and apex of the mandibular bulge.
MNHN AYO 101 ( Fig. 5A, B View FIG ) reveals that the angular process of M. altiplanicum n. sp. is more prominent, with dorsal and ventral margins nearly parallel, than the relatively short and stout process of M. americanum . The medial surface of the angular region bears a well-developed fossa for insertion of the medial pterygoid muscle. A low convexity, lying parallel to the posteroventral margin of the angular process, reflects the depth of the fossa.
The post-molariform part of the dentary is more completely preserved in MNHN BOL V 3294 ( Fig. 5C, D View FIG ). It also indicates that the angular process and insertion for the medial pterygoid were more prominent and deeper, respectively, than in M. americanum . The anterior margin of the ascending ramus rises almost vertically lateral to the distal part of m4, and in lateral view covers part of this tooth, as occurs in M. americanum , M. medinae and M. sundti Philippi, 1893 . In E. laurillardi , however, the anterior margin inclines slightly posteriorly and most of m4 is visible in lateral view ( Hoffstetter 1952).
The condyle resembles that of M. americanum . Its articular surface is transversely elongated and convex. Its medial half is rounded, approximately semicircular, and its lateral half is nearly elliptical and connected to the lateral surface of the ascending ramus by a poorly defined crest.
Lower dentition. All molariforms ( Fig. 4A, B View FIG ) are broken below the level of the alveolar mar- gins, although parts of m1 and m2 are preserved above the margins. The molariforms are quadrangular in section. The transverse axis of m1 lies obliquely, from mesiolingual to distobuccal, to the longitudinal axis of the dentary. m2 is mesiodistally shorter buccally than lingually, and its mesial surface is parallel to the distal surface of m1. The mesial and distal surfaces of m3 are slightly convex, and more nearly perpendicular to the longitudinal axis of the horizontal ramus than those of m1 and m2. The mesial surface of m4 is shorter than the distal surface of m3, and slightly more convex. Its buccal and lingual surfaces con- verge distally. The distal surface of m4 of MNHN AYO 101 has suffered considerable damage, but the alveolar wall of m4 of MNHN BOL V 3294 is very rounded. As in M. lundi seijoi ( Kraglievich 1931) , m1 and m2 are implanted obliquely in the dentary, and incline posteriorly, but m4 is vertical.
Axial Skeleton
Atlas. The altas of the holotype, missing only a part of the right wing, differs in minor details from that of M. americanum . Measurements are given in Table 2. In anterior or posterior view the neural canal is clearly more compressed dorsoventrally than in the Pleistocene species, and the ventral arch is proportionately much wider mediolaterally but shorter anteroposteriorly. Its neural surface bears the fovea dentis along its median length. In E. laurillardi the fovea varies between occupying the whole length and the posterior two thirds of the ventral arch. The ventral tubercle is poorly developed. In anterior view, the anterior articular facets curve more strongly toward the mid-line. The second intervertebral foramen, normally for the passage of blood vessels and the second cervical or spinal nerve, may be formed between the atlas anteriorly and the axis posteriorly. This occurs in MNHN AYO 101. In posterodorsal view of the atlas, this passage forms a smooth, transverse trough between the root of the dorsal arch and the anteromedial wall of the facet for the axis. The trough leads medially into the neural canal; laterally it is bounded by the medial margin of the transverse foramen. However, a second condition may occur, as in MNHN POM 57, where the trough is enclosed by an osseous bridge between the posterodorsal margin of the dorsal arch and the dorsomedial margin of the facet for the axis. This short, transverse canal is similar to the trough in arrangement, probably included the structures that passed between the interverbral foramen, and creates two additional foramina: one opening medially, and the second opening laterally dorsal to the transverse foramen.
Two other foramina are present in M. altiplanicum n. sp. The intervertebral foramen (= lateral vertebral foramen) is a short, transverse canal, which passes into the neural canal. The alar foramen passes dorsoventrally through the anteromedial part of the wing. Whereas the alar and transverse foramina (although better described as canals) become confluent within the lateral mass of the atlas and emerge ventrally upon the atlantal fossa through a common opening in M. americanum , they remain separated in M. altiplanicum n. sp. Hoffstetter (1952) stated that the atlas of E. laurillardi bore the normal four foramina: the alar, transverse, and the anterior and posterior intervertebrals, and apparently considered this as being the normal condition in this species and M. americanum . However, the presence of the posterior intervertebral is variable in E. laurillardi (cf. ROM 4219) and M. altiplanicum n. sp., and presumably also in M. americanum . Such variation apparently also occurs in Megalonyx , and in some specimens the foramen is present only on one side ( McDonald 1977).
The wing is proportionately much larger than in M. americanum . Its dorsal surface bears a welldefined fossa or depression (probably for insertion of the recti capitis muscles) that extends anterior to the anterior intervertebral foramen to reach the level of the anterior margin of the dorsal arch. In M. americanum it does not extend beyond the foramen. Further, the wing in M. altiplanicum n. sp. is distinctly defined anteriorly by an emargination, produced by a lateral prolongation of the dorsolateral margin of the anterior articular facet, between the wing and the facet. In M. americanum the margin of the wing curves without interruption into the dorsolateral margin of the articular facet.
As in M. americanum and other ground sloths, the atlas bears three separate articular facets for the axis: two lateral; the third, median and ventral, articulates with the dens of the axis. The lateral facets are contiguous ventrally with that for the dens in E. laurillardi and E. eomigrans De Iuliis & Cartelle, 1999 ; this condition is apomorphic for Eremotherium ( Cartelle 1992; De Iuliis & Cartelle 1999).
Thoracic vertebrae. The holotype preserves two consecutive thoracic vertebrae and the dorsal arch of the following vertebra. A parapophyseal articular surface lies on the anterior surface of the base of the dorsal arch of all preserved vertebrae. Their centra lack foveae costales. Thus the ribs of the two complete vertebrae articulate only with the parapophyses (i.e. they are syncephalic ribs). In M. americanum this condition occurs only for the last thoracic. This suggests that the complete vertebrae and the dorsal arch belong to the most posterior part of the thoracic series, and that in M. altiplanicum n. sp. vertebrae other than the final thoracic bear syncephalic ribs.
The second complete thoracic vertebra bears secondary or xenarthral postzygapophyses. The dorsal arch that follows bears parapophyses, and is therefore a thoracic vertebra. Further, it bears xenarthral pre- and postzygapophyses. The xenarthral condition thus is present in the posterior two of the three thoracic vertebrae.
Lumbar vertebrae. Two consecutive arches of MNHN AYO 101 are very similar to those of the lumbar vertebrae of M. americanum . They bear xenarthral pre- and postzygapophyses. The anterior of the two arches bears the intratransversary foramen characteristic of the lumbar vertebrae of Xenarthra ; the second arch is damaged and does not permit recognition of the foramen. A distorted neural arch preserves its right postzygapophysis, which articulates with the prezygapophysis of the anterior of the two arches.
Caudal vertebrae. The four consecutive vertebrae of the holotype resemble, based on the shape of their prominent transverse processes, caudal vertebrae 1 to 6 of M. americanum . A poorly defined tubercle lies sagitally on the anterior margin of the neural arch, anterior to the neural spine, of the first of this series, and resembles in size and position those on caudal vertebrae 1 and 2 of M. americanum . The tubercle is absent in the second vertebra of the series. Therefore, the vertebrae of the holotype are probably caudal vertebrae 2 to 5.
Humerus. Only the distal end of a left humerus (MNHN BOL 3338; Table 2) is known for M. altiplanicum n. sp. It is smaller than that in M. americanum , and the epicondyles are less expand- ed and prominent relative to the trochlear surface.
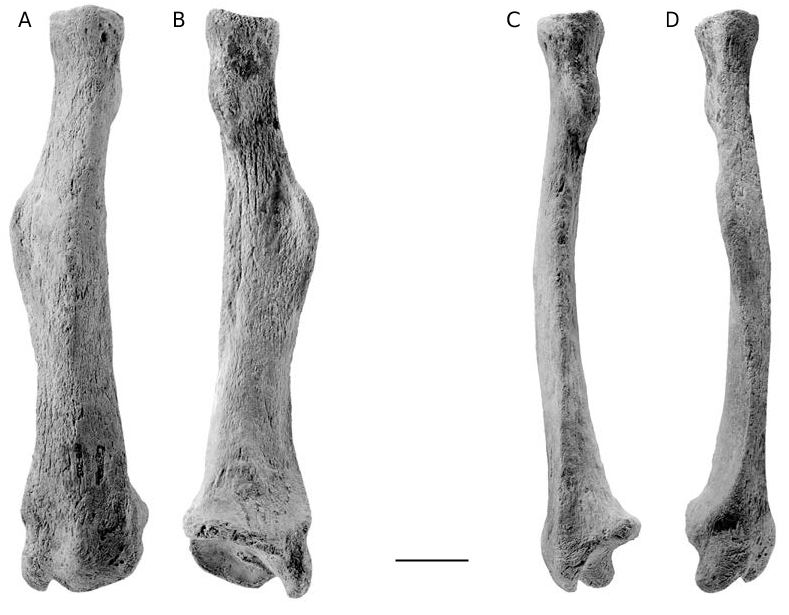
Radius. The left radius (MNHN AYO 203; Fig. 6 View FIG ; Table 2) of M. altiplanicum n. sp. is small- er than but morphologically similar to that of M. americanum . It is also more gracile because its epiphyses are smaller compared to its length. Further, in anterior or posterior views the diaphysis is apparently narrower due to a shorter and less prominent lateral crest, which served largely for the insertion of the pronator teres. The crest is widest within the proximal subterminal quarter of the radius; it lies further distally in M. americanum . There is a pronounced angle between the diaphyseal portions proximal and distal to the crest in M. americanum and M. altiplanicum n. sp., whereas in E. laurillardi , M. tarijense (FMNH P14216) and M. medinae the angle between these portions is less abrupt.
In M. altiplanicum n. sp. the diaphysis is markedly constricted mediolaterally between the crest and the distal end of the radius, and the lateral and medial margins of this part of the diaphysis are subparallel, as occurs in M. americanum . In E. laurillardi , and apparently other megatheriines, the crest lies more distally still, near the midlength of the radius. Further, the diaphysis is constricted slightly just distal to the crest, but gradually widens distally.
The bicipital tuberosity lies more proximally in M. altiplanicum n. sp. and M. americanum than in E. laurillardi . In M. americanum it lies on the medial surface of the radius, in the same transverse plane as the lateral crest, whereas the tuberosity lies more posteriorly, approximately on the medioposterior radial surface in M. altiplanicum n. sp.
Ulna. The proximal part of the right ulna of MNHN AYO 101 is preserved ( Table 2). It is smaller than that of M. americanum and varies morphologically in minor ways. The olecranon process is relatively smaller. The anconeal process is more prominent and distinct. Its margins meet in an acute angle, whereas in M. americanum they form an obtuse angle.
Cuneiform. The cuneiform of megatheriines articulates with the pisiform palmolaterally and the unciform distally. MNHN AYO 150 strongly resembles that of M. americanum , but the facet for the unciform is relatively larger, and that for the pisiform smaller (c. 30% of the maximum width of the cuneiform, as compared to 38% in M. americanum ). However, it is worth noting that Cartelle (1992) reported considerable variation in the shapes and relative sizes of the articular surfaces in E. laurillardi .
Femur. The medial length of the femur ( Table 2) of the holotype ( Fig. 7 View FIG ) is nearly identical to that of the small species Eremotherium sefvei from the Pleistocene of Ulloma, Bolivia (387 mm; De Iuliis & Saint-André 1997), which was probably very similar in body size (cf. infra) to M. altiplanicum n. sp.
The femur of M. altiplanicum n. sp. is smaller than but similar to that of M. americanum . As in the latter, the greater trochanter projects markedly posteriorly to the plane that passes through the head and the distal condyles, so that the bone exhibits strong vertical torsion. In lateral view its lateral margin is sigmoidal. This feature is also present in M. americanum ( Hoffstetter 1952) and M. gallardoi (MACN 5002), but absent in Eremotherium , M. istilarti and M. medinae ( Kraglievich 1925a; Hoffstetter 1952). The last two species, therefore, are not properly considered Megatherium sensu Hoffstetter (1952) . As noted by De Iuliis & Saint-André (1997), torsion of the femur is apparently due to a posterior displacement of the greater trochanter: manipulation of the elements of the hind limb that set the head, greater trochanter, and medial distal condyle in the same plane requires that the long axis of the pes be oriented markedly anteromedially; whereas the pes is oriented anteroposteriorly when the head and distal condyles lie in the same plane.
A
The greater trochanter rises slightly above the crest between the head and greater trochanter. The diameter of the neck is smaller than the maximum diameter of the head, as in M. americanum . The notch for the round ligament is widely triangular, as in M. americanum and E. laurillardi . It is narrow and elongated in E. sefvei and Pyramiodontherium bergi (MLP 2- 66).
The femur of M. altiplanicum n. sp. is relatively narrower than that of M. americanum . As in the latter, the proximal and distal extremities are markedly expanded transversely, the epicondyles prominent, and the lateral and medial margins notably concave, whereas they are more nearly rectilinear in E. laurillardi . In M. medinae the lateral margin is nearly rectilinear, a morphology diagnostic for the species ( Casamiquela & Sepulveda 1974). The less concave margins are probably plesiomorphic states.
The form of the medial condyle is diagnostic for M. altiplanicum n. sp. It extends anterolaterally toward and approaches closely the patellar trochlea. These articular surfaces are in proximity in most megatheriines, but this is due to a medially expanded patellar trochlea (e.g., E. laurillardi ), which is plesiomorphic. The patellar trochlea of M. altiplanicum n. sp. is not medially expanded and thus resembles those of M. americanum and M. gallardoi . In these latter species, the medial articular surface and patellar trochlea are widely separated, particularly in M. gallardoi ( Kraglievich 1925b) . Further, the trochlea does not project anteriorly in M. altiplanicum n. sp. and M. americanum , but does so in E. laurillardi and E. sefvei .
The patellar trochlea is contiguous with the articular surface of the lateral condyle, as in all megatheriines. In M. altiplanicum n. sp., these surfaces are separated by a faint ridge. A similar ridge occurs in some specimens of M. americanum ; a very weak, barely perceptible demarcation may be present in E. laurillardi ( De Iuliis 1996) .
Navicular. A right navicular (MNHN BOL V 3312) from Ayo Ayo is referred to M. altiplanicum n. sp. It is smaller than that of M. americanum and the medial half of the astragalar facet is less concave. This part of the facet is larger than the lateral, convex half, whereas they are subequal in M. americanum . The facets on the anterior surface are arranged as in M. americanum , but the dorsomedial facet is relatively smaller, and the ventral margin of the ventral facet is nearly rectilinear, compared to the convex margin in the Pleistocene species. As with the carpals, Cartelle (1992) reported considerable variation in the shapes and sizes of the articular surfaces of the tarsals in E. laurillardi .
DISCUSSION
The holotype of M. altiplanicum n. sp. represents an adult individual, which had thus attained its definitive size. Two methods for estimating mass suggest that the mass of M. altiplanicum n. sp. may have been been approximately between one quarter to one third that of M. americanum . Fariña et al. (1998) calculated a mass of 6 000 kg for M. americanum based on values obtained from numerous allometric regressions of the dentition, and cranial and postcranial elements. However, these authors obtained a more conservative estimate of 4 000 kg by removing the effect of femur width (which clearly produces a disproportionately high mass estimate in megatheres), a value more similar to the mass obtained by other authors ( Casinos 1996). Using the allometric equation obtained by Janis (1990) based on the lower molar toothrow length of extant ungulates gives a mass of 5 500 kg for M. americanum and 1 746 kg for M. altiplanicum n. sp.
A second method for estimating mass, that of assuming geometric similitude between M. americanum and M. altiplanicum n. sp., results in a lower estimate. Using the specimen measured by Fariña et al. (1998) and a mass of 4 000 kg for M. americanum produces a mass of 977 kg for M. altiplanicum n. sp.; a mass of 6 000 kg for M. americanum produces a mass of 1 465 kg for M. altiplanicum n. sp.
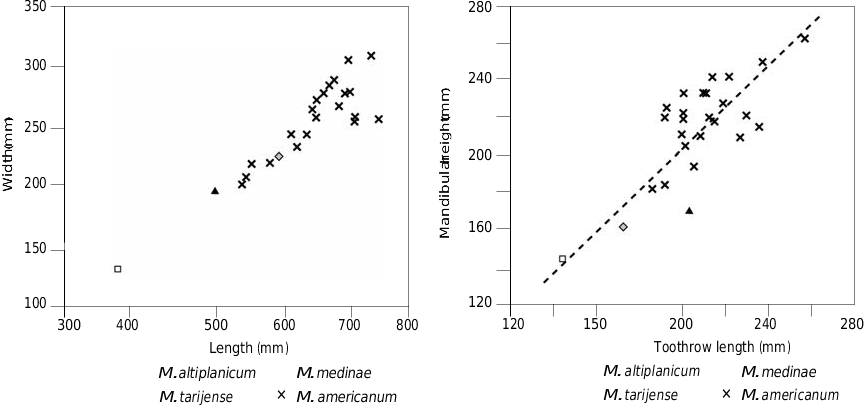
This species is thus considerably smaller than M. americanum and M. gallardoi , and appreciably smaller also than M. medinae , M. istilarti , M. gaudryi and M. tarijense (see Fig. 8). Indeed, it is one of the two smallest post-Miocene megatheriines, the other being Eremotherium sefvei from the Quaternary of Ulloma, Bolivia (De Iuliis & Saint-André 1997).
The toothrow of M. altiplanicum n. sp. (see Fig. 9 View FIG ) is approximately one-third shorter than those of M. medinae and M. tarijense ; these last two species are similar in size ( Casamiquela & Sepulveda 1974). Also, the mandibular toothrow of M. lundi sejoi (see Kraglievich 1931) is 39% longer than that of M. altiplanicum n. sp.
The anterior root of the zygomatic arch lies opposite M1, a position characteristic of the brevirostral (but see De Iuliis 1996) condition described by Cabrera (1928) based on the antemolariform length of the maxilla in some megatheriines. In brevirostral megatheriines, the length between the mesial alveolar wall of M1 and the anterior extremity of the maxilla is just below 17% of the condylomaxillar length, whereas in longirostral megatheriines this ratio is approximately 20%. In the former group, the anterior root of the zygomatic arch lies opposite the middle or distal part of M1, while in the latter it lies at the level of M2. Cabrera (1928) and Paula Couto (1979) stated that divergence of brevirostral and longirostral megatheriines, represented respectively by Megatheridium and Plesiomegatherium , had occurred by the late Miocene. However, a difference of 3% in the ratio of antemolariform maxillar length to condylomaxillar length lies within the range of individual variation in Eremotherium laurillardi ( De Iuliis 1996) . The position of the anterior root of the zygomatic arch lies more posteriorly (i.e. approximately opposite M2) in most megatheriines. However, the polarity of this character is equivocal, but it is worth noting that the root lies in the more posterior position in Megathericulus , which is probably the sister taxon to all other megatheriines, based on its highly mesiodistally compressed molariforms ( De Iuliis 1994).
The palatal morphology of M. altiplanicum n. sp. resembles that of M. medinae and E. laurillardi in that the prealveolar width is nearly equal to the interalveolar width, whereas in M. americanum the prealveolar portion is wider than the interalveolar width. Also, the interalveolar width is narrower that the width of the largest molariform in M. altiplanicum n. sp., M. americanum , M. medinae and Pyramiodontherium bergi , whereas it is equal to or wider than the widest molariform in E. laurillardi , Megatheridium annectens , and Plesiomegatherium halmyrononum ( Cabrera 1928; Hoffstetter 1952).
The femur is strongly twisted in M. americanum and M. altiplanicum n. sp., as is clearly indicated by the markedly sigmoidal curve of the lateral femoral margin. Hoffstetter (1952) considered this feature characteristic of Megatherium . The femur is more nearly flat and its lateral margin nearly rectilinear in Eremotherium , Plesiomegatherium and M. medinae . This morphology is generalized and plesiomorphic for megatheriines, as compared to a femur that is strongly twisted and relatively constricted at its middle (see De Iuliis & Saint-André 1997).
Casamiquela & Sepulveda (1974) considered Eremotherium to be merely a subgenus of
Atlas (holotype, MNHN AYO 101)
Maximum dorsoventral height 56.5 Sagittal length of neural canal 24.0 Maximum dorsoventral height of neural canal 37.5 Maximum transverse width of neural canal 41.0 Dorsoventral height of glenoid fossa 40.0 Maximum length of wing 69.5 Width of wing, from neural canal 54.0 to lateral margin
Humerus (MNHN BOL V 3338)
Maximum distal width 107.2 Width of distal articular surface 70.6 Radius (MNHN AYO 203)
Maximum length 395.0 Maximum proximal diameter 52.5 Minimum proximal diameter 46.2 Maximum distal diameter 99.4 Minimum distal diameter 61.2 Shaft width at level of lateral crest 67.0 Minimum transverse shaft width 42.0 Minimum anteroposterior shaft thickness 27.4 Ulna (MNHN BOL V 3304)
Olecranom process width 119.0 Olecranon process length, tuberosity 88.6 to anconeal process
Width of articular surface of trochlea 103.5 Femur (holotype, MHNH AYO 101)
Lateral length 379.0 Medial length 387.5 Proximal width 203.1 Minimum shaft width 135.0 Maximum diameter of head 94.0 Maximum width between distal tuberosities 196.5 Maximum width between condyles 140.0 Width of patellar trochlea 52.5 Maximum width of medial condyle 64.5 Maximum width of lateral condyle 77.0 and patellar trochlea
Depth of medial condyle 85.0
Megatherium , with M. medinae being a transitional form between primitive species of Eremotherium sensu Hoffstetter (1952) and the more derived species of Megatherium sensu Hoffstetter (1952) . Although this view is probably an oversimplification ( De Iuliis 1996), the well-marked femoral torsion of M. altiplanicum n. sp. avoids misidentification with M. medinae , which possesses the plesiomorphic femoral morphology ( Kraglievich 1931; Hoffstetter 1986).
M. medinae is occasionally referred to as the genus or subgenus Pseudomegatherium Kraglievich, 1931 (“false Megatherium ”); e.g., by Hoffstetter (1986), who stated that at least two as yet undescribed species could possibly be referred to Pseudomegatherium , one from the Pleistocene of Peru, the other from the Quaternary of Ecuador. Casamiquela & Sepulveda (1974), followed by Hoffstetter (1986) and Marshall & Salinas (1991), synonymized M. sundti with M. medinae . However, a femur of an immature individual (PIU M4530) from Ulloma, Bolivia, bears strong resemblance to that of M. americanum , except that the patellar trochlea is expanded medially. This medial expansion occurs in all other megatheriines for which the femur is known, except in M. americanum and M. altiplanicum n. sp. (De Iuliis & Saint-André 1997). Femur PIU M4530 differs sufficiently in general morphology from that of M. medinae to suggest that it belongs to a distinct species. De Iuliis & Saint- André (1997) noted that PIU M4530 probably belonged to M. sundti , described from Ulloma by Philippi (1893), and thus considered this species to be valid. It is worth noting here that Hoffstetter (1986: 224) reported the presence of two megatheriine species from Plio-Pleistocene high Andean mammalian faunas as “ M. sundti = synonym of M. americanum Cuvier ?” However, this passage is misprinted from Hoffstetter’s manuscript for this article, and should read: “ M. sundti Phil. = synonym of M. (Pseudomegatherium) medinae Phil. ; M. americanum Cuvier ?” (Hoffstetter pers. comm. 1991).





| MNHN |
Museum National d'Histoire Naturelle |
| BOL |
University of Cape Town |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
MEGATHERIIDAE Gray, 1821
| Saint-André, Pierre-Antoine & Iuliis, Gerardo De 2001 |
Megatherium altiplanicum
| Saint-André & Iuliis 2001 |
M. altiplanicum
| Saint-André & Iuliis 2001 |
M. altiplanicum
| Saint-André & Iuliis 2001 |
M. altiplanicum
| Saint-André & Iuliis 2001 |
Megatherium
| sensu Hoffstetter 1952 |
Eremotherium
| sensu Hoffstetter 1952 |
Megatherium
| sensu Hoffstetter 1952 |
Megatherium
| sensu Hoffstetter 1952 |
Pseudomegatherium
| Kraglievich 1931 |
Pseudomegatherium
| Kraglievich 1931 |
Plesiomegatherium halmyronomum
| Cabrera 1928 |
Megatheridium annectens
| Cabrera 1928 |
M. medinae
| Philippi 1893 |
M. medinae
| Philippi 1893 |
M. medinae
| Philippi 1893 |
M. medinae
| Philippi 1893 |
M. sundti
| Philippi 1893 |
M. medinae
| Philippi 1893 |
M. medinae
| Philippi 1893 |
M. sundti
| Philippi 1893 |
M. sundti
| Philippi 1893 |
M. sundti Phil.
| Philippi 1893 |
M. (Pseudomegatherium) medinae Phil.
| Philippi 1893 |
Megatherium
| Cuvier 1796 |
M. americanum
| Cuvier 1796 |
M. americanum
| Cuvier 1796 |
M. americanum
| Cuvier 1796 |
M. americanum
| Cuvier 1796 |
M. americanum
| Cuvier 1796 |